El sistema nervioso es uno de los elementos más importantes para nuestra existencia y supervivencia, ya que permite la gestión, organización y funcionamiento del resto de sistemas corporales. Este sistema funciona a través del envío de impulsos electroquímicos con diferentes informaciones y órdenes para las diferentes estructuras que forman parte de nuestro organismo.
Antiguamente se creía que el sistema nervioso era una red continua y sin separación entre elementos, hasta que Ramón y Cajal, por medio de tinturas como la de Golgi, permitió identificar que en realidad está formado por un conjunto de células separadas entre sí: las neuronas. Estas se encuentran separadas por pequeños espacios, pero no dejan de comunicarse entre sí. La conexión existente entre ellas es lo que se conoce como sinapsis.
La sinapsis es una unión intercelular especializada entre neuronas o entre una neurona y una célula efectora, la mayoría de las veces glandular o muscular. En estos contactos se lleva a cabo la transmisión del impulso nervioso, mismo que se inicia con una descarga química que origina una corriente eléctrica en la membrana de la célula presináptica o célula emisora; una vez que este impulso nervioso alcanza el extremo del axón, el cual es la conexión con la otra célula, la propia neurona segrega un tipo de compuestos químicos, los neurotransmisores, que se depositan en el espacio sináptico (espacio intermedio entre esta neurona transmisora y la neurona postsináptica o receptora). Estas sustancias segregadas o neurotransmisores son los encargados de excitar o inhibir la acción de la otra célula llamada célula post sináptica.
Principales componentes
La sinapsis entre dos neuronas, la conexión y vinculación entre ellas que permite que se transmita la información, no es un elemento aislado, sino que está compuesto por tres componentes principales entre los que encontramos parte de ambas neuronas en interrelación: neurona presináptica, espacio sináptico y neurona postsináptica.
Neurona presináptica.- Hace referencia a la neurona que envía la información hacia otra. Esta acción suele llevarse a cabo a través de la emisión de neurotransmisores por parte de las vesículas sinápticas de los botones terminales del final del axón, que a su vez serán recibidos por la membrana de la neurona postsináptica.
Espacio sináptico.- Es el espacio existente entre dos neuronas, generalmente de entre veinte a cuarenta nanómetros. Se trata del espacio en que se produce en sí la transmisión de la información entre neuronas.
Neurona postsináptica.- Parte receptora en la relación entre neuronas. Es la que recibe la información proveniente de la neurona presináptica. Generalmente se trata de las dendritas, aunque dependiendo del tipo de conexión también pueden ser el soma o el axón.
Tipos de sinapsis
Tipos según lo que se transmita
Sinapsis químicas:
Es la más común en el organismo, se transmite de forma química, a través del envío por parte de la neurona presináptica de diferentes neurotransmisores que la neurona postsináptica capta mediante diferentes receptores, cuya acción genera una alteración en forma de potencial excitatorio o inhibitorio postsináptico que puede terminar o no con la generación de un potencial de acción por parte de la neurona postsináptica. Son sinapsis versátiles, puesto que algunas neuronas pueden inhibir la acción de otras dependiendo de qué se active. No existe contacto físico entre ambas neuronas.
Sinapsis eléctricas:
La información se transmite directamente a nivel eléctrico al fluir directamente los iones entre el componente pre y postsináptico. No tienen versatilidad, ya que su actuación no permite que una neurona inhiba la acción de otra. En este tipo de sinapsis existe en realidad un contacto entre neurona pre y postsináptica, a traés de las uniones gap o canales formados por proteínas.
Tipos según el efecto
Sinapsis excitatoria:
Tipo de sinapsis en el que la transmisión de información tiene efectos excitatorios, facilitando que la neurona postsináptica realice un potencial de acción y se continúe la transmisión del mensaje al generar la despolarización de su membrana.
Sinapsis inhibitoria
En este caso, la actuación o activación de este tipo de sinapsis dificulta la aparición de un potencial de acción al hiperpolarizar la célula postsináptica. Se hace más difícil que la información se transmita a través de la neurona postsináptica hacia otras conectadas con ella.
NEUROTRANSMISORES
Los neurotransmisores son sustancias químicas creadas por el cuerpo que transmiten señales (es decir, información) desde una neurona hasta la siguiente a través de unos puntos de contacto llamados sinapsis. Cuando esto ocurre, la sustancia química se libera por las vesículas de la neurona pre-sináptica, atraviesa el espacio sináptico y actúa cambiando el potencial de acción en la neurona post-sináptica.
1. Dopamina
Uno de los más conocidos, que está implicada en las redes cerebrales relacionadas con la motivación y con el comportamiento guiado por recompensas. En este sentido, muchas personas asocian la dopamina con el placer, si bien sería más correcto decir que su actividad depende de la saliencia o del grado de sorpresa que provocan determinados estímulos, también es importante para el movimiento.
2. Adrenalina (o epinefrina)
La adrenalina o epinefrina (término más habitual en inglés que en castellano) es considerada un neurotransmisor cuando actúa en el sistema nervioso, pero posiblemente sean más relevantes sus funciones como hormona -es decir, los efectos que tienen lugar cuando son secretadas en el torrente sanguíneo por las glándulas suprarrenales.
3. Noradrenalina (o norepinefrina)
La noradrenalina también puede ser conceptualizada como un neurotransmisor o como una hormona en función de si ejerce su actividad dentro o fuera del sistema nervioso. Sin embargo, y de modo opuesto a lo que sucede con la adrenalina, en este caso la función de neurotransmisor resulta más relevante que la de hormona.
4. Serotonina (5-HT)
La serotonina también es llamada “5-hidroxitriptamina”; de ahí la abreviatura “5-HT”. En este caso el aminoácido que sirve como precursor no es la tirosina (y en consecuencia la serotonina no forma parte del grupo de las catecolaminas) sino el triptófano, que se puede obtener de alimentos como los huevos, la leche, los cereales integrales o el chocolate, entre otros.
5. Acetilcolina (ACh)
La acetilcolina se deriva de la glucosa que obtenemos a través de la dieta. Entre las funciones del organismo en que participa este neurotransmisor podemos destacar la estimulación (y por tanto la contracción) de las fibras musculares en general y la del cerebro, la producción de saliva, la micción, la erección o la reducción de la frecuencia cardíaca.
6. Glutamato o ácido glutámico (Glu)
El glutamato es el principal neurotransmisor excitatorio del cerebro humano. Está muy extendido por todo este órgano pero, a pesar de la importancia de sus efectos excitatorios, las neuronas que lo utilizan como neurotransmisor son relativamente poco numerosas; por lo general lo emplean con otros objetivos, como la transaminación o la síntesis de proteínas.
7. Ácido gamma-aminobutírico (GABA)
El ácido gamma-aminobutírico, conocido comúnmente por la abreviatura “GABA”, es el neurotransmisor más importante para la inhibición neuronal en el sistema nervioso central, y particularmente en el cerebro. Del GABA depende el tono muscular -de manera que los déficits de este neurotransmisor se relacionan con la hipertonía y la rigidez.
8. Glicina o glicocola (Gly)
El octavo neurotransmisor que hemos incluido en el listado es la glicina, que también tiene efectos inhibitorios en el sistema nervioso central pero, a diferencia del GABA, su actividad es más importante en la médula espinal que en el propio cerebro. La glicina también es muy importante para la síntesis de colágeno, que está presente en la piel y en los huesos.
9. Histamina
Cuando actúa como neurotransmisor en el sistema nervioso, las funciones de la histamina incluyen la regulación del sueño y del estado de alerta, así como la secreción de hormonas por parte del sistema hipotalámico.
La histamina es conocida sobre todo por su papel en las respuestas del sistema inmunitario. Entre otros procesos corporales, este tipo de neurotransmisor se asocia con la inflamación y con las sensaciones de picor.
10. Péptidos opioides endógenos
Existen varios tipos de péptidos opioides que el cerebro produce por él mismo y que son considerados neurotransmisores a causa del modo en que llevan a cabo sus funciones.
El corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre hacia los órganos periféricos. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Los ventrículos aportan la principal fuerza del bombeo que impulsa la sangre.
Una sucesión continuada de contracciones cardíacas denominada ritmicidad cardíaca transmite potenciales de acción por todo el músculo cardíaco y determina su latido rítmico.
Fisiología del músculo cardíaco
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. La duración de la conducción es mucho mayor y las fibras contienen pocas fibrillas contráctiles y presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón.
Anatomía fisiológica del músculo cardíaco
Las fibras musculares cardíacas están dispuestas en un retículo y tienen las miofibrillas típicas con filamentos de actina y miosina.
Músculo cardíaco como sincitio
Los discos intercalados son membranas celulares que separan las células musculares cardíacas individuales entre sí. Es decir, las fibras musculares cardíacas están formadas por muchas células individuales conectadas entre sí en serie y en paralelo. En cada uno de los discos intercalados las membranas celulares se fusionan entre sí de tal manera que forman uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión.
Los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una de ellas se excita el potencial de acción se propaga a todas, propagándose de una célula a otra a través de las interconexiones en enrejado.
El corazón realmente está formado por dos sincitios: el sincitio auricular y el sincitio ventricular. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV). Los potenciales sólo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
Potenciales de acción en el músculo cardíaco
El potencial de acción que se registra en una fibra muscular ventricular es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproxi madamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente + 20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta,seguida de una repolarización súbita.
En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales:
Los mismos canales rápidos de sodio que en el músculo esquelético.
Otra población totalmente distinta de canales lentos de calcio, que también se denominan canales de calcio-sodio, los cuales se abren con mayor lentitud y permanecen abiertos durante varias décimas de segundo.
Inmediatamente después del potencial de acción la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, esto debido al exceso de flujo de entrada de calcio a través de los canales de calcio.
Velocidad de la conducción de las señales en el músculo cardíaco
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de aproximadamente 0,3 a 0,5 m/s.
Periodo refractario del músculo cardíaco
El músculo cardíaco es refractario a la reestimulación durante el potencial de acción. El período refractario del corazón es el intervalo de tiempo, durante el cual un impulso cardíaco normal no puede reexcitar una zona ya excitada de músculo cardíaco. El período refractario normal del ventrículo es de 0,25 a 0,30 s. Hay un período refractario relativo adicional de aproximadamente 0,05 s. El período refractario del músculo auricular es mucho más corto que el de los ventrículos (aproximadamente 0,15 s para las aurículas.
Acoplamiento excitación-contracción: función de los iones calcio y de los túbulos transversos
El término acoplamiento excitación-contracción se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan.
Cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T). Actúan sobre las membranas de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia el sarcoplasma muscular desde el retículo sarcoplásmico.
Además de los iones calcio que se liberan hacia el sarcoplasma desde las cisternas del retículo sarcoplásmico, también difunde una gran cantidad de iones calcio adicionales hacia el sarcoplasma desde los propios túbulos T en el momento del potencial de acción, que abre los canales de calcio dependientes del voltaje a la membrana del túbulo T. El calcio que entra en la célula activa después los canales de liberación de calcio, también denominados canales de receptor de rianodina, en la membrana del retículo sarcoplásmico, para activar la liberación de calcio en el sarcoplasma.
Sin el calcio procedente de los túbulos T la fuerza de la contracción del músculo cardíaco se reduciría de manera considerable porque el retículo sarcoplásmico del músculo cardíaco está peor desarrollado que el del músculo esquelético y no almacena suficiente calcio para generar una contracción completa; en el interior de los túbulos T hay una gran cantidad de mucopolisacáridos que tienen carga negativa y que se unen a una abundante reserva de iones calcio.
La fuerza de la contracción del músculo cardíaco depende en gran medida de la concentración de iones calcio en los líquidos extracelulares. De hecho, un corazón situado en una solución sin calcio dejará rápidamente de latir. La cantidad de iones calcio en el sistema de los túbu- los T (es decir, la disponibilidad de iones calcio para producir la contracción del músculo cardíaco) depende en gran medida de la concentración de iones calcio en el líquido extracelular.
Ciclo cardíaco
Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la gene ración espontánea de un potencial de acción en el nódulo sinusal que está localizado en la pared superolateral de la aurícula derecha. El potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Las aurículas se contraen antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular. Por tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.
Diástole y sístole
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca.
El fonocardiograma es un registro de los ruidos que produce el corazón (principalmente las válvulas cardíacas) durante su función de bombeo.
Efecto de la frecuencia cardíaca en la duración del ciclo cardíaco
Cuando aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye. Cuando aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye. Para una frecuencia cardíaca normal de 72 latidos por minuto, la sístole comprende aproximadamente 0,4 del ciclo cardíaco completo. El corazón que late a una frecuencia muy rápida no permanece relajado el tiempo suficiente para permitir un llenado completo de las cámaras cardíacas antes de la siguiente contracción.
Relación del electrocardiograma con el ciclo cardíaco
El electrocardiograma muestra las ondas P, Q, R, S y T, que son voltajes eléctricos que genera el corazón.
La onda P está producida por la propagación de la despolarización en las aurículas, y es seguida por la contracción auricular.
Las ondas QRS aparecen como consecuencia de la despolarización eléctrica de los ventrículos, que inicia la contracción de los ventrículos; el complejo QRS comienza un poco antes del inicio de la sístole ventricular.
La onda T ventricular representa la fase de repolarización de los ventrículos, se produce antes del final de la contracción ventricular.
Función de las aurículas como bombas de cebado
El 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. Por tanto, las aurículas actúan simplemente como bombas de cebado que aumentan la eficacia del bombeo ventricular. El corazón puede seguir funcionando en la mayor parte de las condiciones.
Cambios de presión en las aurículas: las ondas a, c y v
La onda a está producida por la contracción auricular.
La onda c se produce cuando los ventrículos comienzan a contraerse.
La onda v se produce hacia el final de la contracción ventricular; se debe al flujo lento de sangre hacia las aurículas desde las venas mientras las válvulas AV están cerradas durante la contracción ventricular.
Función de los ventrículos como bombas
Llenado de los ventrículos durante la diástole
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de presión que se ha generado en las aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos. Esto es el periodo de llenado rapido de los ventriculos.
Durante el último tercio de la diástole las aurículas se contraen y aportan un impulso adicional al flujo de entrada de sangre hacia los ventrículos.
Vaciado de los ventrículos durante la sístole
Período de contracción isovolumétrica (isométrica)
Después del comienzo de la contracción ventricular se produce un aumento súbito de presión ventricular, lo que hace que se cierren las válvulas AV. Después son necesarios otros 0,02 a 0,03 s para que el ventrículo acumule una presión suficiente para abrir las válvulas AV semilunares (aórtica y pulmonar) contra las presiones de la aorta y de la arteria pulmonar. Esto se denomina período de contracción isovolumétrica o isométrica.
Periodo de eyección
Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80 mmHg, las presiones ventriculares abren las válvulas semilunares; aproximadamente el 70% del vaciado de la sangre se produce durante el primer tercio del período de eyección y el 30% restante del vaciado durante los dos tercios siguientes. Por tanto, el primer tercio se denomina período de eyección rápida y los dos tercios finales período de eyección lenta.
Período de relajación isovolumétrica (isométrica)
Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones intraventriculares derecha e izquierda disminuyan rápidamente. Durante otros 0,03 a 0,06 s el músculo cardíaco sigue relajándose, aun cuando no se modifica el volu men ventricular, dando lugar al período de relajación isovolumétrica o isométrica. Durante este período las presiones intraventriculares disminuyen rápidamente y regresan a sus bajos valores diastólicos. Después se abren las válvulas AV para comenzar un nuevo ciclo de bombeo ventricular.
Volumen telediastólico, volumen telesistólico y volumen sistólico
Durante la diástole, el llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos hasta aproximadamente 110 a 120 ml. Este volumen se denomina volumen telediastólico. Después, a medida que los ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente 70 ml, lo que se denomina volumen sistólico. El volumen restante que queda en cada uno de los ventrículos, aproximadamente 40 a 50 ml, se denomina volumen telesistólico. La fracción del volumen telediastólico que es propulsada se denomina fracción de eyección.
Función de las válvulas
Válvulas auriculoventriculares
Son las álvulas tricúspide y mitral; impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. Estas válvulas se cierran y abren pasivamente. Casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
Función de los músculos papilares
Los músculos papilares que se unen a los velos de las válvulas AV mediante las cuerdas tendinosas. Los músculos papilares se contraen cuando se contraen las paredes ventriculares, pero no contribuyen al cierre de las válvulas. Tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
Válvula aórtica y de la arteria pulmonar
Las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente; debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV. Los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales.
Curva de presión aórtica
Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica, después la presión del ventrículo aumenta mucho menos rápidamente. La entrada de sangre en las arterias hace que las paredes de las mismas se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Después, al final de la sístole las paredes elásticas de las arterias mantienen una presión elevada en las arterias, incluso durante la diástole. Se produce la denominada incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado. Después de que se haya cerrado la válvula aórtica, la presión en el interior de la aorta disminuye lentamente durante toda la sístole.
Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente ha disminuido hasta aproximadamente 80 mmHg.
Relación de los tonos cardíacos con el bombeo cardíaco
Cuando se contraen los ventrículos primero se oye un ruido que está producido por el cierre de las válvulas AV. La vibración tiene un tono bajo y es relativamente prolongada, y se conoce como el primer tono cardíaco. Cuando se cierran las válvulas aórtica y pulmonar al final de la sístole se oye un golpe seco y rápido porque estas válvulas se cierran rápidamente, y los líquidos circundantes vibran durante un período corto. Este sonido se denomina segundo tono cardíaco.
Generación de trabajo del corazón
El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1min. El trabajo del corazón se utiliza para mover la sangre desde las venas de baja presión hacia las arterias de alta presión. Esto se denomina trabajo volumen-presión o trabajo externo. Además, una pequeña proporción de la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar. Este es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco.
El trabajo del ventrículo izquierdo necesario para crear la energía cinética del flujo sanguíneo es de sólo aproximadamente el 1% del trabajo total del ventrículo y, por tanto, se ignora en el cálculo del trabajo sistólico total.
Análisis gráfico del bombeo ventricular
La curva de presión diastólica se determina llenando el corazón con volúmenes de sangre progresivamente mayores y midiendo la presión diastólica inmediatamente antes de que se produzca la contracción ventricular, que es la presión telediastólica del ventrículo. La curva de presión sistólica se determina registrando la presión sistólica que se alcanza durante la contracción ventricular a cada volumen de llenado.
Hasta los 150 ml a sangre puede fluir con facilidad hacia el ventrículo desde la aurícula, por encima de este valor la presión diastólica ventricular aumenta rápidamente.
Durante la contracción ventricular, la presión «sistólica» aumenta incluso a volúmenes ventriculares bajos y alcanza un máximo a un volumen ventricular de 150 a 170 ml. Los volúmenes elevados los filamentos de actina y de miosina de las fibras musculares cardíacas están tan separados que la fuerza de la contracción de cada una de las fibras cardíacas se hace menos óptima. La presión sistólica máxima del ventrículo izquierdo normal está entre 250 y 300 mmHg. Para el ventrículo derecho normal la presión sistólica máxima está entre 60 y 80 mmHg.
Diagrama volumen-presión durante el ciclo cardíaco; trabajo cardíaco
El bucle denominado diagrama volumen-presión del ciclo cardíaco que sirve para la función normal del ventrículo izquierdo está dividido en 4 fases.
Fase I: Periodo de llenado
Fase II: Periodo de contracción isovolumétrica
Fase III: Periodo de ayección
Fase IV: Periodo de relajación isovolumétrica
Conceptos de precarga y poscarga
Cuando se evalúan las propiedades contráctiles del músculo es importante especificar el grado de tensión del músculo cuando comienza a contraerse, que se denomina precarga, y especificar la carga contra la que el músculo ejerce su fuerza contráctil, que se denomina poscarga.
Para la contracción cardíaca habitualmente se considera que la precarga es la presión telediastólica cuando el ventrículo ya se ha llenado.
Energía química necesaria para la contracción cardíaca: la utilización de oxígeno por el corazón
El músculo cardíaco utiliza energía química para realizar el trabajo de la contracción. Aproximadamente el 70-90% de esta energía procede normalmente del metabolismo oxidativo de los ácidos grasos.
La velocidad del consumo de oxígeno por el miocardio es una medida excelente de la energía química que se libera mientras el corazón realiza su trabajo.
El consumo de oxígeno ha demostrado ser también casi proporcional a la tensión que se produce en el músculo cardíaco durante la contracción multiplicada por la duración de tiempo durante la cual persiste la contracción, denominada índice de tensión-tiempo. Se gasta mucha más energía química a presiones sistólicas normales cuando el ventrículo está dilatado anómalamente debido a que la tensión del músculo cardíaco durante la contracción es proporcional a la presión multiplicada por el diámetro del ventrículo.
La cantidad de energía química necesaria para una cantidad dada de trabajo cardíaco es mayor de lo normal incluso cuando el corazón ya está desfalleciendo.
Regulación del bombeo cardíaco
Regulación intrínseca del bombeo cardíaco: el mecanismo de Frank-Starling
La cantidad de sangre que bombea el corazón cada minuto está determinada, por lo común, casi totalmente por la velocidad del flujo sanguíneo hacia el corazón desde las venas, que se denomina retorno venoso. Esta capacidad intrínseca del corazón de adaptarse a volúmenes crecientes de flujo sanguíneo de entrada se denomina mecanismo de Frank-Starling del corazón. Significa que cuanto más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta.
¿Cuál es la explicación del mecanismo de FrankStarling?
Cuando una cantidad adicional de sangre fluye hacia los ventrículos, el propio músculo cardíaco es distendido hasta una mayor longitud.
La distensión de la pared de la aurícula derecha aumenta directamente la frecuencia cardíaca en un 10-20%; esto también contribuye a aumentar la cantidad de sangre que se bombea cada minuto,
Curvas de función ventricular
Una de las mejores formas de expresar la capacidad funcional de los ventrículos de bombear sangre es mediante la curvas de función ventricular. A medida que aumenta la presión auricular de cada uno de los lados del corazón, el trabajo sistólico de ese lado aumenta hasta que alcanza el límite de la capacidad de bombeo del ventrículo. A medida que aumentan las presiones las aurículas derecha e izquierda, también lo hacen los volúmenes ventriculares por minuto respectivos. Así, las curvas de función ventricular son otra forma de expresar el mecanismo de Frank-Starling del corazón.
Control del corazón por los nervios simpáticos y parasimpáticos
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón. La cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática; el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).
Mecanismos de excitación del corazón por los nervios simpáticos
La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes. Además, aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección.
La nhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado. Cuando la actividad del sistema nervioso simpático disminuye por debajo de lo normal, este fenómeno produce reducción tanto de la frecuencia cardíaca como de la fuerza de la contracción del músculo ventricular, reduciendo de esta manera el nivel de bombeo cardíaco hasta un 30% por debajo de lo normal.
Estimulación parasimpática (vagal) del corazón
La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos. La estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%. Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón. Esto explica el efecto de la estimulación vagal principalmente sobre la reducción de la frecuencia cardíaca.
Efecto de la estimulación simpática y parasimpática sobre la curva de función cardíaca
A cualquier presión auricular derecha dada, el gasto cardíaco aumenta durante el aumento de la estimulación simpática y disminuye durante el aumento de la estimulación parasimpática. Estas modificaciones del gasto que se producen por la estimulación del sistema nervioso autónomo se deben tanto a modi- ficaciones de la frecuencia cardíaca como a modificaciones de la fuerza contráctil del corazón.
Efecto de los iones potasio y calcio sobre la función cardíaca
Efecto de los iones potasio
El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia cardíaca. Grandes cantidades también pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia los ventrículos a través del haz AV. Una concentración elevada de potasio en los líquidos extracelulares reduce el potencial de membrana en reposo de las fibras del músculo cardíaco.
Efecto de los iones calcio
Un exceso de iones calcio hace que el corazón progrese hacia una contracción espástica. El déficit de iones calcio produce flacidez cardíaca y los efectos cardíacos de las concentraciones anormales de calcio raras veces tienen significado clínico.
Efecto de la temperatura sobre la función cardíaca
El aumento de la temperatura corporal produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. La disminución de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta sólo algunos latidos por minuto.
El calor aumenta la permeabilidad de la membrana del músculo cardíaco a los iones que controlan la frecuencia cardíaca, acelerando el proceso de autoexcitación. La fuerza contráctil del corazón con frecuencia aumenta transitoriamente cuando hay un aumento moderado de la temperatura.
El incremento de la carga de presión arterial (hasta un límite) no disminuye el gasto cardíaco
El aumento de la presión arterial en la aorta no reduce el gasto cardíaco hasta que la presión arterial media aumenta por encima de aproximadamente 160 mmHg. El gasto cardíaco está determinado casi totalmente por la facilidad del flujo sanguíneo a través de
los tejidos corporales.
El músculo liso está formado por fibras de 1 a 5 um de diámetro y de sólo 20 a 500 um de longitud. La contracción se da por las mismas fuerzas de atracción entre los filamentos de miosina y actina.
Tipos de músculo liso
El músculo liso se distingue por:
Dimensiones físicas
Organización en fascículos o láminas Respuesta a diferentes tipos de estímulos
Características de la inervación
Función
En general se puede dividir en dos tipos principales:
Músculo liso multiunitario
Formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa. La superficie externa de estas fibras está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí, pero la característica más importante es que cada una de las fibras se puede contraer independientemente de las demás, y su control se ejerce principalmente por señales nerviosas.
Músculo liso unitario o monounitario
Masa de de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra y hacer que las fibras musculares se contraigan simultáneamente.
También se conoce como músculo liso sincitial debido a sus interconexiones sincitiales entre las fibras y como músculo liso visceral porque se encuentra en la pared de la mayor parte de las vísceras del cuerpo.
Mecanismo contráctil en el músculo liso
Base química de la contracción del músculo liso
El músculo liso contiene filamentos tanto de actina como de miosina, pero no contiene el complejo de troponina normal.
El proceso contráctil es activado por los iones calcio, y el trifosfato de adenosina (ATP) se degrada a difosfato de adenosina (ADP) para proporcionar la energía para la contracción.
Base física de la contracción del músculo liso
El músculo liso no tiene la misma disposición estriada de los filamentos de actina y miosina. Grandes números de filamentos de actina unidos a los denominados cuerpos densos, los cuales tienen la misma función que los discos Z. Algunos de estos cuerpos están unidos a la membrana celular y otros están dispersos en el interior de la célula. Algunos de los cuerpos densos de la membrana de células adyacentes están unidos entre sí por puentes proteicos intercelulares. La fuerza de contracción se transmite de unas células a otras a través de estos enlaces. Interpuestos entre los filamentos de actina de la fibra muscular están los filamentos de miosina.
La mayor parte de los filamentos de miosina tiene lo que se denomina puentes cruzados lateropolares, dispuestos de tal manera que los puentes de un lado basculan en una dirección y los del otro lado basculan en la dirección opuesta. Esto permite que la miosina tire de un filamento de actina en una dirección en un lado a la vez que simultáneamente tira de otro filamento de actina en la dirección opuesta en el otro lado, esto permite que las células musculares lisas se contraigan hasta el 80% de su longitud.
Comparación de la contracción del músculo liso con la contracción del músculo estriado
La mayor parte de las contracciones del músculo liso son contracciones tónicas prolongadas, que a veces duran horas o incluso días.
Ciclado lento de los puentes cruzados de miosina
La rapidez del ciclado de los puentes transversos de miosina en el músculo liso es más lenta. La fracción de tiempo que los puentes cruzados permanecen unidos a los filamentos de actina está muy aumentada en el músculo liso. Una posible razón del ciclado lento es que las cabezas de los puentes cruzados tienen una actividad ATPasa mucho menor.
Baja necesidad de energía para mantener la contracción del músculo liso
Para mantener la misma tensión de contracción en el músculo liso es necesario de 1/10 a 1/300 de energía. Sólo es necesaria una molécula de ATP para cada ciclo. La baja utilización de energía por el músculo liso es muy importante para la economía energética global del cuerpo.
Lentitud del inicio de la contracción y relajación del tejido muscular liso total
Un tejido muscular liso típico comienza a contraerse de 50 a 100 ms después de ser excitado, alcanza la contracción completa aproximadamente 0,5 s después, y después la fuerza contráctil disminuye en 1 a 2 segundos más, dando un tiempo total de contracción de 1 a 3 s. La contracción de algunos tipos puede ser tan corta como 0,2 s o tan larga como 30 s. El inicio de la contracciónes es mucho más lento en respuesta a los iones calcio.
La fuerza máxima de contracción muscular es a menudo mayor en el músculo liso que en el músculo esquelético
La fuerza de contracción del músculo liso es mayor, hasta 4 a 6 kg/cm2 de área transversal. Esto se debe al período prolongado de unión de los puentes cruzados de miosina a los filamentos de actina.
El mecanismo de «cerrojo» facilita el mantenimiento prolongado de las contracciones del músculo liso
Una vez que el músculo liso ha generado la contracción máxima, la magnitud de la excitación continuada habitualmente se puede reducir a mucho menos del nivel inicial, a pesar de lo cual el músculo mantiene su fuerza de contracción completa. Además, la energía que se consume para mantener la contracción con frecuencia es minúscula.
Tensión-relajación del músculo liso
Otra característica importante del músculo liso, especialmente del tipo unitario visceral de músculo liso de muchos órganos huecos, es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado. Cuando se produce una reducción súbita de volumen, la presión disminuye drásticamente al principio, aunque después aumenta en un plazo de otros pocos segundos o minutos hasta el nivel original o casi hasta el mismo. Estos fenómenos se denominan tensión-relajación y tensión-relajación inversa.
Regulación de la contracción por los iones calcio
El estímulo que inicia la mayor parte de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular. Puede estar producido por la estimulación nerviosa, por estimulación hormonal, por distensión de la fibra o incluso por cambios del ambiente químico de la fibra. El músculo liso no contiene troponina.
Los iones calcio se combinan con la calmodulina para provocar la activación de la miosina cinasa y fosforilación de la cabeza de miosina
En lugar de la troponina, las células musculares lisas contienen una gran cantidad de otra proteína reguladora denominada calmodulina, la cual activa los puentes cruzados de miosina.
La miosina fosfatasa es importante en la interrupción de la contracción
La inversión de la fosforilación de la cabeza de miosina necesita de la enzima miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y escinde el fosfato de la cadena ligera reguladora. Después se interrumpe el ciclo y finaliza la contracción. El tiempo necesario para la relajación de la contracción muscular está determinado en gran medida por la cantidad de miosina fosfatasa activa en la célula. El número de cabezas unidas a la actina determina la fuerza estática de la contracción.
Control nervioso y hormonal de la contracción del músculo liso
La contracción del músculo liso puede ser estimulada por múltiples tipos de señales:
Señales nerviosas
Estimulación hormonal
Distensión del músculo
Otros estímulos
La membrana del músculo liso contiene muchos tipos de proteínas receptoras que pueden iniciar el proceso contráctil. Otras proteínas receptoras inhiben la contracción del músculo liso.
Uniones neuromusculares del músculo liso
Las ibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares. Generalmente no hacen contacto directo con la membrana de las células de las fibras musculares lisas, sino que forman las denominadas uniones difusas que secretan su sustancia transmisora hacia el recubrimiento de matriz del músculo liso. Las fibras nerviosas con frecuencia inervan solo la capa externa, la excitación muscular viaja desde esta capa externa hacia las capas internas por conducción de los potenciales de acción en la masa muscular o mediante difusión adicional de la sustancia transmisora.
Los axones terminales delgados tiene múltiples varicosidades distribuidas a lo largo de sus ejes. En estos puntos se interrumpen las células de Schwann que rodean a los axones, de modo que se puede secretar la sustancia transmisora a través de las paredes de las varicosidades.
Las vesículas de las terminaciones de las fibras nerviosas autónomas contienen acetilcolina en algunas fibras y noradrenalina en otras, y de manera ocasional también otras sustancias. En el tipo multiunitario del músculo liso, las varicosidades están separadas de la membrana de la célula muscular por tan sólo 20 a 30 nm,Estas uniones se denominan uniones de contacto.
Sustancias transmisoras excitadoras e inhibidoras secretadas en la unión neuromuscular del músculo liso
Las sustancias transmisoras más importantes que secretan los nervios autónomos que inervan el músculo liso son acetilcolina y noradrenalina. La acetilcolina es una sustancia transmisora excitadora de las fibras musculares lisas en algunos órganos y un transmisor inhibidor en el músculo liso de otros órganos. Cuando la acetilcolina excita una fibra, la noradrenalina habitualmente la inhibe y de forma inversa.
Tanto a noradrenalina excitan o inhiben el músculo liso uniéndose en primer lugar a una proteína receptora de la superficie de la membrana de la célula muscular. Algunas de las proteínas receptoras son receptores excitadores, mientras que otras son receptores inhibidores.
Potenciales de membrana y potenciales de acción en el músculo liso
Potenciales de membrana en el músculo liso
El voltaje cuantitativo del potencial de membrana del músculo liso depende de la situación momentánea del músculo. En el estado de reposo normal el potencial intracelular es habitualmente de aproximadamente –50 a –60 mV.
Potenciales de acción en el músculo liso unitario
Los potenciales de acción del músculo liso visceral se producen por potenciales en espiga y por potenciales de acción con meseta.
Potenciales en espiga
Aparecen en la mayor parte de los tipos de músculo liso unitario. La duración de este tipo de potencial de acción es de 10 a 50 ms. Se dan por estimulación eléctrica, por la acción de hormonas sobre el músculo liso, por la acción de sustancias transmisoras procedentes de las fibras nerviosas, por distensión o como consecuencia de su generación espontánea en la propia fibra muscular.
Potenciales de acción con meseta
En lugar de la repolarización rápida de la membrana de la fibra muscular, la repolarización se retrasa durante varios cientos hasta 1.000 ms (1 s). Puede ser responsable de la contracción prolongada que se produce en algunos tipos de músculo liso.
Los canales de calcio son importantes en la generación del potencial de acción del músculo liso
La membrana de la célula muscular lisa tiene muchos más canales de calcio activados por el voltaje, pero pocos canales de sodio. El flujo de iones calcio hacia el interior de la fibra es el principal responsable del potencial de acción. Los canales de calcio se abren muchas veces más lentos que los canales de sodio, y también permanecen abiertos mucho más tiempo.
Los iones calcio actúan directamente sobre el mecanismo contráctil del músculo liso para producir la contracción.
Los potenciales de onda lenta en el músculo liso unitario pueden conducir a la generación espontánea de potenciales de acción
Algunas células musculares lisas son autoexcitadoras. Es decir, los potenciales de acción se originan en las propias células musculares lisas sin ningún estímulo extrínseco. Esto con frecuencia se asocia a un ritmo de ondas lentas básico del potencial de membrana.
Las ondas lentas están producidas por la aparición y desaparición del bombeo de iones positivos hacia el exterior a través de la membrana de la fibra muscular y cuando son lo suficientemente intensas, pueden iniciar potenciales de acción. Cuando el máximo del potencial de la onda lenta negativa en el interior de la membrana celular aumenta en dirección positiva desde –60 hasta aproximadamente –35 mV. Las ondas lentas se denominan ondas marcapasos.
Excitación del músculo liso visceral por distensión muscular
Cuando el músculo liso visceral es distendido lo suficiente, habitualmente se generan potenciales de acción espontáneos, que se deben a una combinación de los potenciales de onda lenta normales y la disminución de la negatividad global del potencial de membrana que produce la propia distensión.
Despolarización del músculo liso multiunitario sin potenciales de acción
Las fibras musculares lisas del músculo liso multiunitario normalmente se contraen sobre todo en respuesta a estímulos nerviosos. Las sustancias transmisoras producen despolarización de la membrana del músculo liso, y esto a su vez produce la contracción.
En las células musculares lisas pequeñas, incluso sin potencial de acción, la despolarización local, denominada potencial de la unión, que produce la propia sustancia transmisora nerviosa se propaga «electrotónicamente» en toda la fibra y es lo único necesario para producir la contracción muscular.
Los efectos de los factores tisulares locales y las hormonas determinan la contracción del músculo liso sin potenciales de acción
Contracción del músculo liso en respuesta a factores químicos tisulares locales
El músculo liso es muy contráctil y responde rápidamente a los cambios de las condiciones químicas locales del líquido intersticial circundante. Un potente sistema de control de retroalimentación local controla el flujo sanguíneo a la zona tisular local. Algunos de los factores de control específicos son los siguientes:
La ausencia de oxígeno en los tejidos locales produce relajación del músculo liso y, por tanto, vasodilatación.
El exceso de anhídrido carbónico produce vasodilatación.
El aumento de la concentración de iones hidrógeno produce vasodilatación.
La adenosina, el ácido láctico, el aumento de los iones potasio, la disminución de la concentración de los iones calcio y el aumento de la temperatura corporal producen vasodilatación local.
Efectos de las hormonas sobre la contracción del músculo liso
Las hormonas circulantes en la sangre afectan en cierto grado a la contracción del músculo liso, y algunas tienen efectos profundos. Entre las más importantes se encuentran la noradrenalina, la adrenalina, la acetilcolina, la angiotensina, la endotelina, la vasopresina, la oxitocina, la serotonina y la histamina.
Una hormona produce contracción del músculo liso cuando la membrana de la célula muscular contiene receptores excitadores activados por hormonas para esa hormona. Por el contrario, la hormona produce inhibición si la membrana contiene receptores inhibidores para ella.
Mecanismos de la excitación o la inhibición del músculo liso por hormonas o por factores tisulares locales
Algunos receptores hormonales de la membrana del músculo liso abren canales iónicos de sodio o de calcio y despolarizan la membrana, al igual que ocurre después de la estimulación nerviosa. Algunas veces la contracción o la inhibición del músculo liso es iniciada por hormonas que no producen directamente ningún cambio en el potencial de membrana. En estos casos la hormona puede activar un receptor de membrana que no abre ningún canal iónico, sino que produce un cambio interno de la fibra muscular.
Para inhibir la contracción se sabe que otros mecanismos activan la enzima adenilato ciclasa o guanilato ciclasa de la membrana celular; las porciones de los receptores que sobresalen hacia el interior de las células están acopladas con estas enzimas, dando lugar a la formación de monofosfato cíclico de adenosina (AMPc) o monofosfato cíclico de guanosina (GMPc), denominados segundos mensajeros.
Origen de los iones calcio que causan la contracción a través de la membrana celular y a partir del retículo sarcoplásmico
Aunque el proceso contráctil del músculo liso es actiado por iones calcio, el origen de estos es diferente. El retículo sarcoplásmico está poco desarrollado en la mayor parte del músculo liso. Por el contrario, la mayoría de los iones calcio que producen la contracción entran en la célula muscular desde el líquido extracelular en el momento del potencial de acción o de otro estímulo.
El tiempo necesario para que se produzca esta difusión es en promedio de entre 200 y 300 ms y se denomina período de latencia antes de que comience la contracción.
Función del retículo sarcoplásmico del músculo liso
Las cavéolas son pequeñas invaginaciones de la membrana celular, serían un análogo rudimentario del sistema de túbulos transversos del músculo esquelético. Se piensa que la transmisión de un potencial de acción hacia las cavéolas excita la liberación de iones calcio desde los túbulos sarcoplásmicos próximos. Mientras más extenso sea el retículo sarcoplásmico de la fibra muscular lisa, más rápidamente se contraerá.
La contracción del músculo liso depende de la concentración extracelular de los iones calcio
Cuando la concentración de iones calcio en el líquido extracelular disminuye a aproximadamente 1/3 a 1/10 de su valor normal, habitualmente se interrumpe la contracción del músculo. La fuerza de la contracción del músculo liso depende mucho de la concentración de iones calcio en el líquido extracelular.
Es necesaria una bomba de calcio para producir la relajación del músculo liso
Para producir la relajación del músculo liso después de que se haya contraído se deben retirar los iones calcio de los líquidos intracelulares. Esta eliminación se consigue mediante una bomba de calcio que bombea iones calcio hacia el exterior de la fibra muscular lisa de nuevo hacia el líquido extracelular o hacia el retículo sarcoplásmico, si está presente.
Las fibras del músculo esquelético están inervadas por fibras nerviosas mielinizadas grandes que se originan en las motoneuronas grandes de las astas anteriores de la médula espinal. Cada terminación nerviosa forma una unión, denominada unión neuromuscular, con la fibra muscular cerca de su punto medio.
Anatomía fisiológica de la unión neuromuscular: la placa motora terminal
La fibra nerviosa forma un complejo de terminaciones nerviosas ramificadas que se invaginan en la superficie de la fibra muscular, pero que permanecen fuera de la membrana plasmática de la misma. Toda la estructura se denomina placa motora terminal. Esta cubierta por una o más células de Schwann que la aíslan de los líquidos circundantes.
La membrana invaginada se denomina gotiera sináptica o valle sináptico y el espacio que hay entre la terminación y la membrana de la fibra se denomina espacio sináptico o hendidura sináptica. Este espacio mide de 20 a 30 nm de anchura. En el fondo de la gotiera hay numerosos pliegues más pequeños de la membrana de la fibra muscular denominados hendiduras subneurales, que aumentan mucho el área superficial en la que puede actuar el transmisor sináptico.
En la terminación axónica hay muchas mitocondrias que proporcionan trifosfato de adenosina (ATP) que es la fuente de energía que se utiliza para la síntesis de la acetilcolina. En el espacio sináptico hay grandes cantidades de la enzima acetilcolinesterasa.
Secreción de acetilcolina por las terminaciones nerviosas
Cuando un impulso nervioso llega a la unión neuromuscular, se liberan aproximadamente 125 vesículas de acetilcolina desde las terminaciones hacia el espacio sináptico. En la superficie interna de la membrana neural hay barras densas lineales.
A ambos lados de cada una de estas barras densas hay partículas proteicas que penetran en la membrana neural; son canales de calcio activados por el voltaje.
Los iones calcio ejercen una influencia de atracción sobre las vesículas de acetilcolina, desplazándolas hacia la membrana neural adyacente a las barras densas.
Las vesículas se fusionan con la membrana neural y vacían su acetilcolina hacia el espacio sináptico mediante el proceso de exocitosis.
Efecto de la acetilcolina sobre la membrana de la fibra muscular postsináptica para abrir canales iónicos
Los canales iónicos activados por acetilcolina están localizados casi totalmente mente cerca de las aberturas de las hendiduras subneurales, donde se libera la acetilcolina hacia el espacio sináptico. El complejo está formado por cinco subunidades proteicas, dos proteínas alfa y una proteína beta, una delta y una gamma, que atraviesan la membrana, y están dispuestas en círculo para formar un canal tubular.
El canal activado por acetilcolina tiene un diámetro de aproximadamente 0,65 nm, que es lo suficientemente grande como para permitir que los iones positivos importantes se muevan con facilidad a través de la abertura. Fluyen muchos más iones sodio a través de los canales activados por acetilcolina que de cualquier otro tipo.
El principal efecto de la apertura de los canales activados por la acetilcolina es permitir que grandes cantidades de iones sodio entren al interior de la fibra, desplazando con ellos grandes números de cargas positivas. Esto genera un cambio de potencial positivo local en la membrana de la fibra muscular, denominado potencial de la placa terminal. A su vez, este potencial de la placa terminal inicia un potencial de acción que se propaga a lo largo de la membrana muscular y de esta manera produce la contracción muscular.
Destrucción por la acetilcolinesterasa de la acetilcolina liberada
La acetilcolina sigue activando los receptores de acetilcolina mientras persista en el espacio. Sin embargo, se elimina rápidamente por dos medios:
La mayor parte de la acetilcolina es destruida por la enzima acetilcolinesterasa.
Una pequeña cantidad de acetilcolina difunde hacia el exterior del espacio sináptico y ya no está disponible para actuar sobre la membrana de la fibra muscular.
La rápida eliminación de la acetilcolina impide la reexcitación muscular continuada después de que la fibra muscular se haya recuperado de su potencial de acción inicial.
Potencial de la placa terminal y excitación de la fibra muscular esquelética
La rápida entrada de iones sodio en la fibra muscular hace que el potencial eléctrico en el interior de la fibra en la zona local de la placa terminal aumente en dirección positiva hasta 50 a 75 mV, generando un potencial local denominado potencial de la placa terminal.
El curare es un fármaco que bloquea la acción activadora de la acetilcolina sobre los canales de acetilcolina compitiendo con los puntos del receptor de acetilcolina.
La toxina botulínicaes un veneno bacteriano que reduce la magnitud de la liberación de acetilcolina por las terminaciones nerviosas.
Factor de seguridad para la transmisión en la unión neuromuscular; fatiga de la unión
Cada impulso que llega a la unión neuromuscular produce un potencial de la placa terminal aproximadamente tres veces mayor que el necesario para estimular la fibra nerviosa. Por tanto, se dice que la unión neuromuscular normal tiene un elevado factor de seguridad. La estimulación de la fibra nerviosa disminuye tanto el número de vesículas de acetilcolina que los impulsos no pueden pasar hacia la fibra nerviosa. Esto se denomina fatiga de la unión neuromuscular.
Biología molecular de la formación y liberación de acetilcolina
Con cada potencial de acción habitualmente se produce la lisis de aproximadamente 125 vesículas. Posteriormente, después de algunos milisegundos, la acetilcolina es escindida por la acetilcolinesterasa en ion de acetato y colina, y la colina se reabsorbe activamente en la terminación neural para ser reutilizada para formar de nuevo acetilcolina. Esta secuencia de acontecimientos se produce en un período de 5 a 10 ms.
Fármacos que potencian o bloquean la transmisión en la unión neuromuscular
Fármacos que estimulan la fibra muscular por su acción similar a la acetilcolina
La metacolina, carbacol y nicotina, tienen el mismo efecto sobre la fibra muscular que la acetilcolina. Estos fármacos no son destruidos por la colinesterasa, o son destruidos muy lentamente . Actúan produciendo zonas localizadas de despolarización de la membrana de la fibra muscular en la placa motora terminal donde están localizados los receptores de acetilcolina.
Fármacos que estimulan la unión neuromuscular median- te la inactivación de la acetilcolinesterasa
La neostigmina, fisostigmina y fluorofosfato de diisopropilo, inactivan la acetilcolinesterasa de las sinapsis de modo que ya no pueda hidrolizar la acetilcolina. Por tanto, con cada impulso nervioso sucesivo se acumula una cantidad adicional de acetilcolina, que estimula repetitivamente la fibra muscular. Esto produce espasmo muscular. Neostigmina y fisostigmina se combinan con la acetilcolinesterasa para inactivar la acetilcolinesterasa durante hasta varias horas, en cambio el fluorofosfato de diisopropilo inactiva la acetilcolinesterasa durante semanas, lo que hace que sea un tóxico particularmente letal.
Fármacos que bloquean la transmisión en la unión neuromuscular
Los fármacos curariformes puede impedir el paso de los impulsos desde la terminación nerviosa hacia el músculo.
Miastenia grave que causa parálisis muscular
La miastenia grave produce parálisis muscular debido a que las uniones neuromusculares no pueden transmitir suficientes señales desde las fibras nerviosas a las fibras musculares. En cuanto a su patogenia, en la sangre se han detectado anticuerpos dirigidos frente a los receptores de acetilcolina.
Es una enfermedad autoinmunitaria en la que los pacientes han desarrollado anticuerpos que bloquean o destruyen sus propios receptores de acetilcolina en la unión neuromuscular postsináptica.
Potencial de acció muscular
Algunos de los aspectos cuantitativos de los potenciales musculares son los siguientes:
Potencial de membrana en reposo: aproximadamente –80 a –90 mV en las fibras esqueléticas.
Duración del potencial de acción: 1 a 5 ms en el músculo esquelético.
Velocidad de conducción: 3 a 5 m/s, aproximadamente 1/13 de la velocidad de conducción de las fibras nerviosas mielinizadas grandes que excitan al músculo esquelético.
Propagación del potencial de acción al interior de la fibra muscular a través de los túbulos transversos
Para producir una contracción muscular máxima la corriente debe penetrar en las zonas profundas de la fibra muscular hasta la vecindad de las miofibrillas individuales. Esto se consigue mediante la transmisión de los potenciales de acción a lo largo de los túbulos transversos (túbulos T). Los potenciales de acción de los túbulos T producen liberación de iones calcio en el interior de la fibra muscular en la vecindad inmediata de las miofibrillas, y estos iones calcio a su vez producen la contracción. Este proceso global se denomina acoplamiento excitacióncontracción.
Acoplamiento excitación-contracción
Sistema de túbulos transversos-retículo sarcoplásmico
Los túbulos T son pequeños y siguen un trayecto transversal a las miofibrillas; se ramifican entre ellos y forman planos completos de de túbulos T que se entrelazan entre todas las miofibrillas individuales. Además, donde los túbulos T se originan en la membrana celular, están abiertos hacia el exterior de la fibra muscular. Por tanto, se comunican con el líquido extracelular que rodea la fibra muscular y contienen líquido extracelular en su luz. Se podría decir que los túbulos T son realmente extensiones internas de la membrana celular. Las corrientes eléctricas que rodean a estos túbulos T producen la contracción muscular.
Hay un retículo sarcoplásmico formado por dos partes principales:
Grandes cavidades denominadas cisternas terminales, que están junto a los túbulos T.
Túbulos longitudinales largos que rodean todas las superficies de las miofibrillas que se están contrayendo.
Liberación de iones calcio por el retículo sarcoplásmico
En el interior de sus túbulos vesiculares hay un exceso de iones calcio a una concentración elevada y muchos son liberados desde cada una de las vesículas cuando se produce un potencial de acción en el túbulo T adyacente.
El potencial de acción del túbulo T genera un flujo de corriente hacia las cisternas del retículo sarcoplásmico en su punto de contacto con el túbulo T. Cuando el potencial de acción alcanza al túbulo T, el cambio de voltaje es detectado por receptores de dihidropiridina que están ligados a canales de liberación de calcio, también denominados canales receptores de rianodina, en las cisternas reticulares sarcoplásmicas adyacentes. La activación de los receptores de dihidropiridina provoca la apertura de los canales de liberación de calcio en las cisternas, así como en sus túbulos longitudinales anexos.
Bomba de calcio para retirar los iones calcio del líquido miofibrilar después de que se haya producido la contracción
La contracción muscular continúa mientras los iones calcio permanezcan a una concentración elevada. Una bomba de calcio que actúa continuamente y que está localizada en las paredes del retículo sarcoplásmico bombea iones calcio desde las miofibrillas de nuevo hacia los túbulos sarcoplásmicos.
Además, en el interior del retículo hay una proteína denominada calsecuestrina, que puede unirse a hasta 40 veces más calcio.
Pulso excitador de los iones calcio
El complejo troponina-tropomiosina mantiene inhibidos los filamentos de actina y mantiene el estado relajado del músculo. Por el contrario, la excitación completa del sistema del túbulo T y del retículo sarcoplásmico da lugar a una liberación de iones calcio suficiente como para aumentar la concentración en el líquido miofibrilar.
La duración total de este pulso de calcio en la fibra muscular esquelética normal dura aproximadamente 1/20 de segundo y durante éste se produce la contracción muscular.
Todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 mm y se extienden a lo largo de toda la longitud del músculo. Excepto por aproximadamente el 2% de las fibras, todas están habitualmente inervadas sólo por una terminación nerviosa.
El sarcolema es una fina membrana que envuelve a una fibra musculoesquelética
El sarcolema está formado por una membrana plasmática, y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno.
Las miofibrillas están formadas por filamentos de actina y miosina
Cada fibra muscular contiene varios cientos a varios miles de miofibrillas. Cada una de estas miofibrillas está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí.
Los filamentos de miosina y de actina se interdigitan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas. Las bandas claras contienen sólo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada. Pequeñas proyecciones se originan en los lados de los filamentos de miosina y se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción.
Los extremos de los filamentos de actina están unidos al denominado disco Z, el cual atraviesa las miofibrillas y también pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular.
La porción de la miofibrilla (o de la fibra muscular entera) que está entre dos discos Z sucesivos se denomina sarcómero. Cuando la fibra muscular está contraída la longitud del sarcómero es de aproximadamente 2 um.
Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina
La relación de yuxtaposición entre los filamentos de miosina y de actina se consigue con titina, la cual es una de las mayores moléculas proteicas del cuerpo. Es filamentosa y muy elástica. Actúan como armazón que mantiene en su posición los filamentos de miosina y de actina, de modo que funcione la maquinaria contráctil del sarcómero. Un extremo de la molécula de titina es elástico y está unido al disco Z. La otra parte de la molécula de titina la une al grueso filamento de miosina.
El sarcoplasma es el fluido intracelular entre las miofibrillas
Los espacios entre las miofibrillas están llenos de un líquido intracelular denominado sarcoplasma, que contiene grandes cantidades de potasio, magnesio y fosfato, además de múltiples enzimas proteicas. También hay muchas mitocondrias que están dispuestas paralelas a las miofibrillas. Las mitocondrias proporcionan a las miofibrillas en contracción grandes cantidades de energía en forma de trifosfato de adenosina (ATP).
El retículo sarcoplásmico es un retículo endoplásmico especializado de músculo esquelético
En el sarcoplasma que rodea a las miofibrillas hay un extenso retículo denominado retículo sarcoplásmico, cuya organización especial que es muy importante para controlar la contracción muscular.
Mecanismo general de la contracción muscular
Mecanismo molecular de la contracción muscular
Mecanismo de deslizamiento de los filamentos de la contracción muscular
En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí. Por el contrario, en el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión. Además, los discos Z han sido traccionados por los filamentos de actina hasta los extremos de los filamentos de miosina. Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos.
Este fenómeno está producido por las fuerzas que se generan por la inte racción de los puentes cruzados que van desde los filamentos de miosina a los filamentos de actina. Para que se realice el proceso contráctil se necesita de energía, misma que procede de los enlaces de alta energía de la molécula de ATP, que es degradada a difosfato de adenosina (ADP) para liberarla.
Características moleculares de los filamentos contráctiles:
Los filamentos de miosina están compuestos por múltiples moléculas de miosina.
Cada una de las moléculas de miosina tiene un peso molecular de aproximadamente 480000. La molécula de miosina está formada por seis cadenas polipeptídicas, dos cadenas pesadas, cada una con un peso molecular de aproximadamente 200000, y cuatro cadenas ligeras, que tienen un peso molecular de aproximadamente 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Un extremo de cada una de estas cadenas se pliega bilateralmente para formar una estructura polipeptídica globular denominada cabeza de la miosina. Las cuatro cadenas ligeras también forman parte de la cabeza de la miosina, dos en cada cabeza. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular.
El filamento de miosina está formado por 200 o más moléculas individuales de miosina. Parte del cuerpo de cada una de las moléculas de miosina se prolonga hacia la región lateral junto a la cabeza, formando de esta manera un brazo que separa la cabeza del cuerpo, como se muestra en la figura. Los brazos y las cabezas que protruyen se denominan en conjunto puentes cruzados.
Cada puente cruzado es flexible en dos puntos denominados bisagras. Los brazos articulados permiten que las cabezas se separen del cuerpo del filamento de miosina o que se aproximen al mismo. Las cabezas articuladas, a su vez, participan en el proceso real de contracción. La longitud total de los filamentos de miosina es uniforme, casi exactamente 1,6 um.
El propio filamento de miosina está enrollado de modo que cada par sucesivo de puentes cruzados está desplazado en sentido axial 120° respecto al par previo.
Actividad ATPasa de la cabeza de miosina
La cabeza de miosina actúa como una enzima ATPasa, lo cual permite que la cabeza escinda el ATP y que utilice la energía procedente del enlace fosfato de alta energía del ATP para aportar energía al proceso de la contracción.
Los filamentos de actina están formados por actina, tropomiosina y troponina
El esqueleto del filamento de actina es una molécula de la proteína F-actina bicatenaria. Las dos hélices están enroscadas en una hélice. Cada una de las hebras de la doble hélice de F-actina está formada por moléculas de G-actina polimerizadas. A cada una de estas moléculas de G-actina se le une una molécula de ADP. Se piensa que estas moléculas de ADP son los puntos activos de los filamentos de actina con los que interactúan los puentes cruzados de los filamentos de miosina para producir la contracción muscular. Los puntos activos de las dos hebras de F-actina están escalonados, lo que permite que haya un punto activo en toda la longitud del filamento de actina cada 2,7 nm. Cada uno de los filamentos de actina tiene una longitud de aproximadamente 1 mm. Las bases de los filamentos de actina se anclan fuertemente en los discos Z.
Moléculas de tropomiosina
El filamento de actina también contiene otra proteína, la tropomiosina, que están enrolladas en espiral alrededor de los lados de la hélice de F-actina. En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
Troponina y su función en la contracción muscular
Unidas de manera intermitente a lo largo de los lados de las moléculas de tropomiosina están las troponinas. Se trata de complejos de tres subunidades proteicas unidas entre sí de manera laxa.
Una de las subunidades (troponina I) tiene una gran afinidad por la actina, otra (troponina T) por la tropomiosina y la tercera (troponina C) por los iones calcio. Se piensa que este complejo une la tropomiosina a la actina. Se piensa que la intensa afinidad de la troponina por los iones calcio inicia el proceso de la contracción.
Interacción de un filamento de miosina, dos filamentos de actina y los iones calcio para producir la contracción
Inhibición del filamento de actina por el complejo troponina-tropomiosina; activación por los iones calcio
Se piensa que los puntos activos del filamento de actina normal del músculo relajado son inhibidos o cubiertos físicamente por el complejo troponina-tropomiosina.
Antes de que se produzca la contracción, se debe inhibir el efecto bloqueante del complejo troponina-tropomiosina. Cuando hay grandes cantidades de iones calcio, se inhibe el propio efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina.
La relación normal entre el complejo troponina-tropomiosina y la actina es alterada por los iones calcio, dando lugar a una nueva situación que lleva a la contracción.
Interacción entre el filamento de actina «activado» y los puentes cruzados de miosina: teoría de la «cremallera» de la contracción
Tan pronto como el filamento de actina es activado por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y de algún modo esto hace que se produzca la contracción. Una hipótesis para la que hay datos considerables es la teoría de la cremallera o teoría del trinquete de la contracción.
Se ha propuesto que cuando una cabeza se une a un punto activo, esta unión produce simultáneamente cambios profundos en las fuerzas intramoleculares entre la cabeza y el brazo de este puente cruzado. La nueva alineación de las fuerzas hace que la cabeza se desplace hacia el brazo y que arrastre con ella al filamento de actina. Este desplazamiento de la cabeza se denomina golpe activo. Inmediatamente después del desplazamiento, la cabeza se separa automáticamente del punto activo.
Las cabezas de los puentes cruzados se incurvan hacia atrás y hacia delante y paso a paso recorren el filamento de actina, desplazando los extremos de dos filamentos de actina sucesivos hacia el centro del filamento de miosina. Cada uno de los puentes cruzados actúa independientemente de todos los demás, uniéndose y tirando en un ciclo repetido continuo. Por tanto, cuanto mayor sea el número de puentes cruzados que estén en contacto con el fila- mento de actina en un momento dado, mayor será la fuerza de contracción.
ATP como fuente de energía para la contracción: fenómenos químicos en el movimiento de las cabezas de miosina
Durante el proceso de contracción se escinden grandes cantidades de ATP para formar ADP; cuanto mayor sea la magnitud del trabajo que realiza el músculo, mayor será la cantidad de ATP que se escinde, lo que se denomina efecto Fenn.
El proceso se realiza una y otra vez hasta que los filamentos de actina han desplazado la membrana Z hasta los extremos de los filamentos de miosina o hasta que la carga que se ejerce sobre el músculo se hace demasiado grande como para que se produzca una tracción adicional.
El efecto de la cantidad de superposición de los filamentos de actina y miosina determina la tensión desarrollada por el músculo en contracción.
Efecto de la longitud muscular sobre la fuerza de contracción en el músculo intacto entero
El músculo entero tiene una gran cantidad de tejido conjuntivo; además, los sarcómeros de diferentes partes del músculo no siempre se contraen la misma magnitud.
Cuando el músculo está en su longitud normal en reposo, que corresponde a una longitud del sarcómero de aproximadamente 2 mm, se contrae con una fuerza de contracción próxima a la fuerza máxima cuando es activado. Sin embargo, el aumento de la tensión que se produce durante la contracción, denominado tensión activa, se reduce a medida que el músculo es distendido más allá de su longitud normal, es decir, hasta una longitud del sarcómero mayor de aproximadamente 2,2 um.
Relación de la velocidad de contracción con la carga
Un músculo esquelético se contrae rápidamente cuando lo hace frente a una carga nula, hasta un estado de contracción completa en aproximadamente 0,1 s para un músculo medio. Cuando se aplican cargas, la velocidad de la contracción se hace cada vez más lenta a medida que aumenta la carga, es decir, cuando la carga ha aumentado hasta la fuerza máxima que puede ejercer el músculo, la velocidad de contracción se hace cero y no se produce ninguna contracción, a pesar de la activación de la fibra muscular. La fuerza neta de que se dispone para producir la velocidad de acortamiento está reducida de manera proporcional.
Energética de la contracción muscular
Generación de trabajo durante la contracción muscular
Cuando un músculo se contrae contra una carga realiza un trabajo. En términos matemáticos el trabajo se define mediante la siguiente ecuación:
T = C × D
donde T es el trabajo generado, C es la carga y D es la distancia del movimiento que se opone a la carga. La energía necesaria para realizar el trabajo procede de las reacciones químicas de las células musculares durante la contracción.
Fuentes de energía para la contracción muscular
La contracción muscular depende de la energía que aporta el ATP, aunque son necesarias cantidades pequeñas para:
Bombear iones calcio desde el sarcoplasma hacia el interior del retículo sarcoplásmico después de que haya finalizado la contracción.
Para bombear iones sodio y potasio a través de la membrana de la fibra muscular para mantener un entorno iónico adecuado para la propagación de los potenciales de acción de la fibra muscular.
La contracción de ATP en la fibra muscular es suficiente para mantener la contracción completa durante sólo 1 a 2 s como máximo. El ATP se escinde para formar ADP, que transfiere la energía de la molécula de ATP a la maquinaria contráctil de la fibra muscular.
La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que se escinde inmediatamente y la energía que se libera produce el enlace de un nuevo ion fosfato al ADP para reconstituir el ATP. La energía combinada del ATP y de la fosfocreatina almacenados en el músculo es capaz de producir una contracción muscular máxima durante sólo 5 a 8 s.
La segunda fuente importante de energía es la glucólisis del glucógeno que se ha almacenado previamente en las células musculares. La escisión enzimática rápida del glucógeno en ácido pirúvico y ácido láctico libera energía que se utiliza para convertir el ADP en ATP, las reacciones glucolíticas se pueden producir incluso en ausencia de oxígeno.
La tercera y última fuente de energía es el metabolismo oxidativo. Esto supone combinar oxígeno con los productos finales de la glucólisis y con otros diversos nutrientes celulares para liberar ATP. Más del 95% de toda la energía que utilizan los músculos para la contracción sostenida a largo plazo procede de esta fuente. Los nutrientes que se consumen son carbohidratos, grasas y proteínas.
Características de la contracción de todo el músculo
Muchas características de la contracción muscular se pueden demostrar desencadenando espasmos musculares únicos.
Contracción isométrica frente a isotónica
Se dice que la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción.
Las características de la contracción isotónica dependen de la carga contra la que se contrae el músculo, así como de la inercia de la carga. Sin embargo, el sistema isométrico registra estrictamente los cambios de la fuerza de la propia contracción muscular.
Características de los espasmos isométricos que se registran en diferentes múscul
El cuerpo humano tiene músculos esqueléticos de muchos tamaños. Las fibras pueden ser tan pequeñas como de 10 mm de diámetro o tan grandes como de 80 mm. Finalmente, la energética de la contracción muscular varía considerablemente de un músculo a otro.
Fibras musculares rápidas frente a lentas
Todos los músculos del cuerpo están formados por una mezcla de las denominadas fibras musculares rápidas y lentas, con otras fibras intermedias entre estos dos extremos. Las diferencias entre estos dos tipos de fibras son las siguientes:
Fibras lentas (tipo I, músculo rojo):
Fibras más pequeñas.
Inervadas por fibras nerviosas pequeñas.
Vascularización y capilares más extensos para aportar cantidades adicionales de oxígeno.
Números muy elevados de mitocondrias para mantener niveles elevados de metabolismo oxidativo.
Las fibras contienen grandes cantidades de mioglobina, misma que se combina con el oxígeno y lo almacena hasta que sea necesario, además de darle al músculo lento un aspecto rojizo y el nombre de músculo rojo.
Fibras rápidas (tipo II, músculo blanco):
Fibras grandes para obtener una gran fuerza de contracción.
Retículo sarcoplásmico extenso para una liberación rápida de iones calcio para iniciar la contracción.
Grandes cantidades de enzimas glucolíticas para la liberación rápida de energía por el proceso glucolítico.
Vascularización menos extensa porque el metabolismo oxidativo tiene una importancia secundaria.
Menos mitocondrias, también porque el metabolismo oxidativo es secundario.
Un déficit de mioglobina roja en el músculo rápido le da el nombre de músculo blanco.
Mecánica de la contracción del músculo esquelético:
Unidad motora: todas las fibras musculares inervadas por una única fibra nerviosa
Todas las fibras musculares que son inervadas por una única fibra nerviosa se denominan unidad motora. En general, los músculos pequeños que reaccionan rápidamente y cuyo control debe ser exacto tienen más fibras nerviosas para menos fibras musculares. Los músculos que no precisan un control fino pueden tener varios centenares de fibras musculares en una unidad motora. Una cifra promedio sería de aproximadamente 80 a 100 fibras musculares por unidad motora. Las fibras musculares de todas las unidades motoras se superponen a otras unidades motoras en microfascículos de 3 a 15 fibras. Esta interdigitación permite que las unidades motoras separadas se contraigan cooperando entre sí.
Sumación significa la adición de los espasmos individuales para aumentar la intensidad de la contracción muscular global, se produce de dos maneras:
Aumentando el número de unidades motoras que se contraen de manera simultánea, lo que se denomina sumación de fibras múltiples.
Aumentando la frecuencia de la contracción, lo que se denomina sumación de frecuencia y que puede producir tetanización.
Sumación de fibras múltiples
las unidades motoras de mayor tamaño con frecuencia tienen una fuerza contráctil hasta 50 veces mayor que las unidades más pequeñas. Esto se denomina principio de tamaño. Es importante porque permite que se produzcan gradaciones de la fuerza muscular durante la contracción débil en escalones pequeños, mientras que los escalones se hacen cada vez mayores cuando son necesarias grandes cantidades de fuerza. La causa de este principio de tamaño es que las unidades motoras más pequeñas son activadas por fibras nerviosas motoras pequeñas, y que las motoneuronas pequeñas de la médula espinal son más excitables que las grandes, de modo que naturalmente se excitan antes.
Las diferentes unidades motoras son activadas de manera sincrónica por la médula espinal, de modo que la contracción se alterna entre las unidades motoras de manera secuencial.
Sumación de frecuencia y tetanización
A medida que aumenta la frecuencia, se llega a un punto en el que cada nueva contracción se produce antes de que haya finalizado la anterior; la fuerza total de la contracción aumenta progresivamente al aumentar la frecuencia. Cuando la frecuencia alcanza un nivel crítico, las contracciones sucesivas finalmente se hacen tan rápidas que se fusionan entre sí, y la contracción del músculo entero parece ser completamente suave y continua, esto se denomina tetanización.
Máxima fuerza de contracción
La máxima fuerza de contracción tetánica de un músculo que funciona a una longitud muscular normal es en promedio de entre 3 y 4 kg por centímetro cuadrado de músculo.
Cambios de la fuerza muscular al inicio de la contracción: el efecto de la escalera (Treppe)
Cuando un músculo comienza a contraerse después de un período de reposo prolongado, su fuerza de contracción inicial puede ser tan pequeña, es decir, la fuerza de la contracción aumenta hasta una meseta, un fenómeno que se denomina efecto de la escalera o Treppe. Se piensa que está producido principalmente por el aumento de los iones calcio en el citosol debido a la liberación de cada vez más iones desde el retículo sarcoplásmico con cada potencial de acción muscular sucesivo y la incapacidad del sarcoplasma de recapturar inmediatamente los iones.
Tono del músculo esquelético
Incluso cuando los músculos están en reposo habitualmente hay una cierta cantidad de tensión, que se denomina tono muscular, el cual se debe totalmente a impulsos nerviosos de baja frecuencia que proceden de la médula espinal. Estos, a su vez, están controlados en parte por señales que se transmiten desde el encéfalo a las motoneuronas adecuadas del asta anterior de la médula espinal y en parte por señales que se originan en los husos musculares.
Fatiga muscular
La contracción prolongada e intensa de un músculo da lugar al conocido estado de fatiga muscular, ésta aumenta en una proporción casi directa a la velocidad de depleción del glucógeno del músculo. Por tanto, la fatiga se debe principalmente a la incapacidad de los procesos contráctiles y metabólicos de las fibras musculares de continuar generando el mismo trabajo. La interrupción del flujo sanguíneo a través de un músculo que se está contra yendo da lugar a una fatiga muscular casi completa en un plazo de 1 a 2 min debido a la pérdida de aporte de nutrientes, especialmente de oxígeno.
Sistemas de palanca del cuerpo
El análisis de los sistemas de palanca del cuerpo depende del conocimiento de:
El punto de la inserción muscular
Su distancia desde el fulcro de la palanca
La longitud del brazo de la palanca
La posición de la palanca
Hay muchos tipos diferentes de músculo; algunos son largos y se contraen una distancia larga, y algunos son cortos pero tienen áreas transversales grandes y pueden proporcionar una fuerza de contracción extrema en distancias pequeñas. El estudio de los diferentes tipos de músculos, de los sistemas de palanca y de sus movimientos se denomina cinesiología.
Colocación de una parte del cuerpo por la contracción de los músculos agonistas y antagonistas de lados opuestos de una articulación: «coactivación» de los músculos antagonistas
Todos los movimientos del cuerpo están producidos por la contracción simultánea de músculos agonistas y antagonistas de lados opuestos de las articulaciones. Esto se denomina coactivación de los músculos agonistas; y antagonistas y está controlada por los centros de control motor del encéfalo y de la médula espinal. La posición de cada una de las partes separadas del cuerpo está determinada por los grados relativos de contracción de los músculos agonistas y antagonistas.
Remodelado del músculo para adaptarse a la función
Todos los músculos del cuerpo se modelan continuamente para adaptarse a las funciones que deben realizar. Se altera su diámetro, su longitud, su fuerza y su vascularización, e incluso se alteran, al menos ligeramente, los tipos de fibras musculares. Se produce en pocas semanas.
Hipertrofia y atrofia muscular
Cuando se produce un aumento de la masa total de un músculo se denomina hipertrofiamuscular. Cuando disminuye, el proceso se denomina atrofia muscular. Prácticamente toda la hipertrofia muscular se debe a un aumento del número de filamentos de actina y miosina en cada fibra muscular, dando lugar a aumento de tamaño de las fibras musculares individuales; esto se denomina hipertrofia de las fibras. La hipertrofia aparece en un grado mucho mayor cuando el músculo está sometido a carga durante el proceso contráctil.
La velocidad de síntesis de las proteínas contráctiles del músculo es mucho mayor cuando se está produciendo la hipertrofia y también se produce un aumento de los sistemas enzimáticos que proporcionan energía.
Cuando un músculo no se utiliza durante muchas semanas, la velocidad de degradación de las proteínas contráctiles es mucho más rápida que la velocidad de sustitución. Por tanto, se produce atrofia muscular. La ruta que parece importar en buena parte para la degradación proteica en un músculo que experimenta atrofia es la ruta de ubicuitina-proteasoma dependiente del ATP.
Ajuste de la longitud muscular
Otro tipo de hipertrofia se produce cuando los músculos son distendidos hasta una longitud mayor de lo normal. Esto hace que se añadan nuevos sarcómeros en los extremos de las fibras musculares, donde se unen a los tendones. Cuando un músculo permanece acortado a una longitud menor que su longitud normal de manera continua, los sarcómeros de los extremos de las fibras musculares pueden llegar realmente a desaparecer.
Hiperplasia de las fibras musculares
Cuando se da un aumento del número de fibras se denomina hiperplasia de las fibras. Cuando aparece, el mecanismo es la división lineal de fibras que estaban previamente aumentadas de tamaño.
Efectos de la denervación muscular
Cuando un músculo pierde su inervación, ya no recibe las señales contráctiles que son necesarias para mantener el tamaño muscular normal. Por tanto, la atrofia comienza casi inmediatamente. Si la inervación del músculo se restaura rápidamente, se puede producir la recuperación completa de la función en un plazo tan pequeño como 3 meses.
En la fase final de la atrofia por denervación, la mayor parte de las fibras musculares son destruidas y sustituidas por tejido fibroso y adiposo.
El tejido fibroso que sustituye a las fibras musculares durante la atrofia por denervación también tiende a seguir acortándose durante muchos meses, lo que se denomina contractura.
Recuperación de la contracción muscular en la poliomielitis: aparición de macrounidades motoras
LAs fibras musculares paralizadas. Esto da lugar a unidades motoras de gran tamaño denominadas macrounidades motoras, que pueden contener hasta cinco veces el número normal de fibras musculares para cada neurona que procede de la médula espinal.
Rigidez cadavérica
Varias horas después de la muerte, todos los músculos del cuerpo entran en un estado de contractura denominado rigidez cadavérica. Esta rigidez se debe a la pérdida de todo el ATP. El músculo permanece rígido hasta que las proteínas del músculo se deterioran aproximadamente 15 a 25 h después.
Algunas células como las células nerviosas y musculares, generan impulsos electroquímicos rápidamente cambiantes en sus membranas, y estos impulsos se utilizan para transmitir señales a través de las membranas de los nervios y de los músculos. En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células.
Física básica de los potenciales de membrana
Potenciales de membrana provocados por difusión
«Potencial de difusión» producido por una diferencia de concentración iónica a los dos lados de la membrana
Una diferencia de concentración de iones a través de una membrana puede, en condiciones adecuadas, crear un potencial de membrana. Y la diferencia de potencial entre el interior y el exterior se denomina potencial de difusión.
En la fibra nerviosa normal del mamífero la diferencia de potencial para el potasio necesaria es de aproximadamente 94 mV, con negatividad en el interior de la membrana de la fibra; en cambio para el sodio el potencial es de aproximadamente 61 mV positivos en el interior de la fibra.
Muchos de los rápidos cambios de los potenciales de membrana que se observan durante la transmisión de los impulsos nerviosos y musculares se deben a la aparición de estos potenciales de difusión rápidamente cambiantes.
Relación del potencial de difusión con la diferencia de concentración: potencial de Nernst
El nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana se denomina potencial de Nernst para ese ion. La magnitud viene determinada por el cociente de las concentraciones de ese ion específico en los dos lados de la membrana.
Se puede utilizar la ecuación de Nernst, para calcular el potencial de Nernst para cualquier ion univalente a la temperatura corporal normal (37 °C). El signo del potencial es positivo (+) si el ion que difunde desde el interior hacia el exterior es un ion negativo, y es negativo (–) si el ion es positivo.
Cálculo del potencial de difusión cuando la membrana es permeable a varios iones diferentes
Cuando una membrana es permeable a varios iones diferentes, el potencial de difusión que se genera depende de tres factores:
La polaridad de la carga eléctrica de cada uno de los iones
La permeabilidad de la membrana a cada uno de los iones
Las concentraciones de los respectivos iones en el interior y en el exterior de la membrana
La ecuación de Goldman o ecuación de Goldman-Hodgkin-Katz, da el potencial de membrana calculado en el interior de la membrana cuando participan dos iones positivos univalentes, sodio (Na+) y potasio (K+), y un ion negativo univalente, cloruro (Cl–).
Esta ecuación es importante ya que:
Los iones sodio, potasio y cloruro son los iones más importantes que participan en la generación de los potenciales de membrana en las fibras nerviosas y musculares. El gradiente de concentración de cada uno de estos iones a través de la membrana ayuda a determinar el voltaje del potencial de membrana.
El grado de importancia de cada uno de los iones en la determinación del voltaje es proporcional a la permeabilidad de la membrana para ese ion particular.
Un gradiente positivo de concentración iónica desde el interior de la membrana hacia el exterior produce electronegatividad en el interior de la membrana. Se produce el efecto contrario cuando hay un gradiente de un ion negativo.
La permeabilidad de los canales de sodio y de potasio experimenta cambios rápidos durante la transmisión de un impulso nervioso, mientras que la permeabilidad de los canales de cloruro no se modifica mucho durante este proceso.
Medición del potencial de membrana
Una pipeta pequeña llena de una solución de electrólitos y se inserta en la membrana celular hasta el interior de la fibra. Después se coloca otro electrodo, denominado «electrodo indiferente», en el líquido extracelular, y se mide la diferencia de potencial entre el interior y exterior de la fibra utilizando un voltímetro adecuado, el cual es un aparato electrónico muy sofisticado que puede medir voltajes pequeños a pesar de la resistencia muy elevada al flujo eléctrico a través de la punta de la micropipeta. Para registrar los cambios rápidos del potencial de membrana durante la transmisión de los impulsos nerviosos el microelectrodo se conecta a un osciloscopio.
Siempre que el electrodo esté fuera de la membrana del nervio el potencial que se registra es cero, que es el potencial del líquido extracelular. Después, a medida que el electrodo de registro atraviesa la zona de cambio de voltaje en la membrana celular (denominada capa de dipolo eléctrico) el potencial disminuye bruscamente hasta –90 mV. Al moverse a través del interior de la fibra el potencial permanece en un nivel estable de –90 mV, aunque vuelve a cero en el momento en el que atraviesa la membrana en el lado opuesto de la fibra.
Para generar un potencial negativo en el interior de la membrana sólo se debe transportar hacia fuera un número suficiente de iones positivos para generar la capa de dipolo eléctrico en la propia membrana. Todos los demás iones del interior de la fibra nerviosa pueden ser positivos o negativos.
Potencial de membrana en reposo de los nervios
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV, es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.
Transporte activo de los iones sodio y potasio a través de la membrana: la bomba sodio-potasio (Na+-K+)
La bomba de sodio-potasio se trata de una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior, dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular. También genera grandes gradientes de concentración para el sodio y el potasio a través de la membrana nerviosa en reposo. Estos gradientes son los siguientes:
Na+ (exterior): 142 mEq/l
Na+ (interior): 14 mEq/l
K+ (exterior): 4 mEq/l
K+ (interior): 140 mEq/l
Los cocientes de estos dos iones respectivos desde el interior al exterior son:
Na+ (interior) / Na+ (exterior)= 0,1
K+ (interior) / K+ (exterior)= 35
Fuga de potasio y de sodio a través de la membrana nerviosa
Existe una proteína del canal denominada dominio de poros en tándem, canal de potasio o canal de fuga de potasio (K+), en la membrana nerviosa a través de la que pueden escapar iones potasio incluso en una célula en reposo, también pueden dejar que se pierdan algunos iones de sodio pero los canales son mucho más permeables al potasio que al sodio y esta diferencia de permeabilidad es un factor clave para determinar el nivel del potencial de membrana en reposo normal.
Origen del potencial de membrana en reposo normal
Los factores más importantes que establecen el potencial de membrana en reposo normal de –90 mV son los siguientes:
Contribución del potencial de difusión de potasio
Debido al elevado cociente de los iones potasio entre el interior y el exterior, si los iones potasio fueran el único factor que genera el potencial en reposo, el potencial en reposo en el interior de la fibra sería igual a –94 mV.
Contribución de la difusión de sodio a través de la membrana nerviosa
El cociente de los iones sodio desde el interior hasta el exterior de la membrana es de 0,1 y da un potencial de Nernst de +61 mV. El potencial de Nernst para la difusión de potasio es de -94 mV. Utilizando la ecuación de Goldman se ve que si la membrana es muy permeable al potasio pero sólo ligeramente permeable al sodio, es lógico que la difusión del potasio contribuya mucho más al potencial de membrana que la difusión del sodio. Utilizando este valor en la ecuación de Goldman se obtiene un potencial en el interior de la membrana de –86 mV.
Contribución de la bomba Na+-K+
El hecho de que se bombeen más iones sodio hacia el exterior que iones potasio hacia el interior genera un grado adicional de negatividad (aproximadamente –4 mV más) en el interior además del que se puede explicar por la difusión de manera aislada.
Los potenciales de difusión aislados que produce la difusión del sodio y del potasio darían un potencial de membrana de aproximadamente –86 mV, esto más los –4 mV adicionales al potencial de membrana por la acción continua de la bomba de Na+-K+ electrógena, genera un potencial neto de membrana de –90 mV.
Potencial de acción nervioso
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo.
Las fases del potencial de acción son las siguientes:
Fase de reposo
Fase de despolarización
Fase de repolarización
Canales de sodio y potasio activados por el voltaje
Tienen una función adicional a la bomba de Na+-K+ y de los canales de fuga K+.
Canal de sodio activado por el voltaje: activación e inactivación del canal
Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. En reposo normal la compuerta de activación está cerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra.
Activación del canal de sodio
Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo alcanza un voltaje (–70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta, esto se denomina estado activado.
Inactivación del canal de sodio
El mismo aumento de voltaje que abre la compuerta de activación también cierra la compuerta de inactivación, pero lo hace alguas diezmilésimas de segundo después de que se abra la compuesta de activación. En este punto el potencial de membrana comienza a recuperarse de nuevo hacia el estado de membrana en reposo, lo que es el proceso de repolarización.
La compuerta de inactivación no se abre de nuevo hasta que el potencial de membrana se normaliza o casi a valores de reposo. Por tanto, en general el canal de sodio no se puede abrir de nuevo sin que antes se repolarice la fibra nerviosa.
Canal de potasio activado por el voltaje y su activación
Durante el estado de reposo la compuerta del canal de potasio está cerrada. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce una apertura conformacional de la compuerta y permite el aumento de la difusión de potasio hacia fuera a través del canal. Se abren al mismo tiempo que están comenzando a cerrarse los canales de sodio debido a su inactivación. Por tanto, la disminución de la entrada de sodio hacia la célula y el aumento simultáneo de la salida de potasio desde la célula se combinan para acelerar el proceso de repolarización.
Resumen de los fenómenos que causan el potencial de acción
Durante el estado de reposo la conductancia a los iones potasio es 50 a 100 veces mayor que la conductancia a los iones sodio, pero al inicio del potencial de acción se activan instantáneamente los canales de sodio y dan lugar a un aumento de la conductancia al sodio de 5.000 veces.
Durante la primera porción del potencial de acción el cociente de la conductancia al sodio respecto a la del potasio aumenta más de 1.000 veces; esto es lo que hace que el potencial de membrana se haga positivo al inicio del potencial de acción. Después empiezan a cerrarse los canales de sodio y empiezan a abrirse los canales de potasio, de modo que el cociente de conductancias se desplaza más a favor de la elevada conductancia al potasio con una baja conductancia al sodio. En consecuencia, el potencial de acción vuelve rápidamente a su nivel basal.
Funciones de otros iones durante el potencial de acción
Iones con carga negativa (aniones) no difusibles en el interior del axón nervioso
En el interior del axón hay muchos iones de carga negativa que no pueden atravesar los canales de la membrana. Incluyen los aniones de las moléculas proteicas y de muchos compuestos de fosfato orgánicos, compuestos de sulfato y similares; son responsables de la carga negativa en el interior de la fibra cuando hay un déficit neto de iones potasio de carga positiva y de otros iones positivos.
Iones calcio
El calcio coopera con el sodio o actúa en su lugar en algunas células para producir la mayor parte del potencial de acción. La bomba de potasio transporta iones calcio desde el interior hacia el exterior de la membrana celular (o hacia el interior del retículo endoplásmico de la célula), creando un gradiente de ion calcio de aproximadamente 10.000 veces. Hay canales de calcio activados por el voltaje y existe un enorme gradiente de difusión para el flujo pasivo de iones calcio a las células.
Una función importante de los canales de iones calcio activados por voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. No obstante, la activación de los canales de calcio es lenta, y precisa hasta 10 a 20 veces más tiempo para su activación que los canales de sodio. Por este motivo, a menudo se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción.
En algunos tipos de músculo liso los potenciales de acción están producidos casi totalmente por la activación de los canales lentos de calcio.
Aumento de la permeabilidad de los canales de sodio cuando hay déficit de iones calcio
Cuando hay déficit de iones calcio, los canales de sodio se activan (abren) por un pequeño aumento del potencial de membrana desde su nivel normal, muy negativo. Es necesario que la concentración del ion calcio disminuya sólo un 50% por debajo de su concentración normal para que se produzca la descarga espontánea en algunos nervios periféricos, produciendo con frecuencia «tetania» muscular.
Inicio del potencial de acción
Un ciclo de retroalimentación positiva abre los canales de sodio
Si algún episodio produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje. Esto permite la entrada rápida de iones sodio, lo que produce una elevación adicional del potencial de membrana y abre aún más canales de sodio activados por el voltaje y permite que se produzca una mayor entrada de iones sodio hacia el interior de la fibra. Este proceso es un círculo vicioso de retroalimentación positiva que continúa hasta que se han activado todos los canales de sodio activados por el voltaje. Posteriormente, el aumento del potencial de membrana produce cierre de los canales de sodio y apertura de los canales de potasio, y pronto finaliza el potencial de acción.
Umbral para el inicio del potencial de acción
No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva. Un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90 mV hasta aproximadamente –65 mV habitualmente da lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.
Propagación del potencial de acción
Un potencial de acción que se desencadena en cualquier punto de una membrana excitable habitualmente excita porciones adyacentes de la membrana, dando lugar a la propagación del potencial de acción a lo largo de la membrana.
Las cargas eléctricas positivas son desplazadas por la difusión hacia dentro de iones sodio a través de la membrana despolarizada y posteriormente a lo largo de varios milímetros en ambos sentidos a lo largo del núcleo del axón. Estas cargas positivas aumentan el voltaje a lo largo de una distancia de 1 a 3 mm a lo largo de la gran fibra mielinizada hasta un valor superior al umbral del voltaje para iniciar el potencial de acción. Por tanto, los canales de sodio de estas nuevas zonas se abren inmediatamente y se produce una propagación explosiva del potencial de acción. Estas zonas recién despolarizadas producen a su vez más circuitos locales de flujo de corriente en zonas más lejanas de la membrana, produciendo una despolarización progresivamente creciente. Esta transmisión del proceso de despolarización a lo largo de una fibra nerviosa muscular se denomina impulso nervioso o muscular.
Dirección de la propagación
Una membrana excitable no tiene una dirección de propagación única, sino que el potencial de acción viaja en todas las direcciones alejándose del estímulo hasta que se ha despolarizado toda la membrana.
Principio del todo o nada
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son., este principio se aplica a todos los tejidos excitables normales.
para que se produzca la propagación continuada de un impulso, en todo momento el cociente del potencial de acción respecto al umbral de excitación debe ser mayor de 1. Este requisito de mayor de 1 se denomina factor de seguridad para la propagación.
Restablecimiento de los gradientes iónicos de sodio y potasio tras completarse los potenciales de acción: la importancia del metabolismo de la energía
La propagación de cada potencial de acción a lo largo de una fibra nerviosa reduce ligeramente las diferencias de concentración de sodio y de potasio en el interior y en el exterior de la membrana. Se pueden transmitir entre 100.000 y 50 millones de impulsos por las grandes fibras nerviosas de gran tamaño antes de que las diferencias de concentración alcancen el punto de que se interrumpa la conducción del potencial de acción. Con el tiempo se hace necesario restablecer las diferencias de las concentraciones de membrana de sodio y de potasio, es decir, los iones sodio que han difundido hacia el interior de la célula durante los potenciales de acción y los iones potasio que han difundido hacia el exterior deben volver a su estado original por la bomba Na+K+. Como esta bomba precisa energía para esta operación, esta «recarga» de la fibra nerviosa es un proceso metabólico activo que utiliza la energía que procede del sistema energético del trifosfato de adenosina (ATP) de la célula.
Una característica especial de la bomba Na+K+ATPasa es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular.
Meseta en algunos potenciales de acción
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máxi mo del potencial de espiga durante muchos milisegundos, esto prolonga el período de despolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas, en las dura hasta 0,2 a 0,3 s.
En el proceso de despolarización del músculo cardíaco la apertura de los canales rápidos origina la porción en espiga del potencial de acción, mientras que la apertura prolongada de los canales lentos de calciosodio principalmente permite la entrada de iones calcio en la fibra, lo que es responsable en buena medida también de la porción de meseta del potencial de acción.
Los canales de potasio activados por el voltaje tienen una apertura más lenta de lo habitual, y con frecuencia no se abren mucho hasta el final de la meseta.
Ritmicidad de algunos tejidos excitables: descarga repetitiva
Las descragas repetitivas aparecen en mayormente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central y casi todos los demás tejidos excitables pueden descargar de manera repetitiva si se reduce lo suficiente el umbral de estimulación de las células del tejido.
Proceso de reexcitación necesario para la ritmicidad espontánea
Para que se produzca ritmicidad espontánea la membrana debe ser lo suficientemente permeable a los iones sodio (o a los iones calcio y sodio a través de los canales lentos de calciosodio) como para permitir la despolarización automática de la membrana.
El flujo aumentado de salida de iones potasio desplaza grandes cantidades de cargas positivas hacia el exterior de la membrana, dejando en el interior de la fibra una negatividad mucho mayor de lo que se produciría de otra manera. Esto continúa durante aproximadamente un segundo después de que haya finalizado el po tencial de acción anterior, acercando de esta manera el potencial de membrana al potencial de Nernst del potasio. Este es un estado denominado hiperpolarización. Siempre que exista este estado no se producirá autoexcitación. Pero la conductancia aumentada para el potasio (y el estado de hiperpolarización) desaparece gradualmente, después de que haya finalizado el potencial de acción, lo que permite que el potencial de membrana aumente de nuevo hasta el umbral de excitación.
Características especiales de la transmisión de señales en los troncos nerviosos
Fibras nerviosas mielinizadas y no mielinizadas
Las fibras grandes son mielinizadas y las pequeñas no mielinizadas. Un tronco nervioso medio contiene aproximadamente el doble de fibras no mielinizadas que mielinizadas.
El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier, el cual es una pequeña zona no aislada en la unión entre 2 células de Schwann sucesivas a lo largo del axón en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón.
Las células de Schwann depositan la vaina de mielina alrededor del axón.
Conducción saltatoria en las fibras mielinizadas de un nódulo a otro
Los potenciales de acción se producen solo en los nódulos y se conducen desde un nódulo a otro, esto es lo que se conoce como conducción saltatoria, es decir, el impulso nervioso recorre a saltos la fibra.
La conducción saltatoria es útil porque aumenta la velocidad de la transmisión nerviosa en las fibras mielinizadas hasta 5 a 50 veces; conserva la energía para el axón porque sólo se despolarizan los nódulos, permitiendo una pérdida de iones tal vez 100 veces menor de lo que sería necesario de otra manera y precisa poco metabolismo para restablecer las diferencias de concentración de sodio y de potasio a través de la membrana después de una serie de impulsos nerviosos; además, el excelente aislamiento que ofrece la membrana de mielina y la disminución de 50 veces de la capacitancia de la membrana permiten que se produzca la repolarización con poca transferencia de iones.
Velocidad de conducción en las fibras nerviosas
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan solo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (un valor superior a la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
Excitación: el proceso de generación del potencial de acción
Cualquier factor que haga que los iones sodio comiencen a difundir hacia el interior a través de la membrana en un número suficiente puede desencadenar la apertura regenerativa automática de los canales de sodio. Esto se puede deber a un trastorno mecánico de la membrana, a los efectos químicos sobre la membrana o al paso de electricidad a través de la membrana.
Excitación de una fibra nerviosa por un electrodo metálico cargado negativamente
La corriente negativa desde el electrodo reduce el voltaje del exterior de la membrana hasta un valor negativo más próximo al voltaje del potencial negativo del interior de la fibra. Esto reduce el voltaje eléctrico a través de la membrana y permite que se abran los canales de sodio, lo que da lugar a un potencial de acción. Por el contrario, en el electrodo positivo la inyección de cargas positivas sobre el exterior de la membrana nerviosa aumenta la diferencia de voltaje a través de la membrana en lugar de reducirla. Esto produce un estado de hiperpolarización, que realmente reduce la excitabilidad de la fibra en lugar de producir un potencial de acción.
Umbral de excitación y potenciales locales agudos
Un estímulo eléctrico negativo débil puede no ser capaz de excitar una fibra. Sin embargo, cuando aumenta el voltaje del estímulo se llega a un punto en el que se produce la excitación.
Los cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generar un potencial de acción, se denominan potenciales subliminales agudos.
Cuando el potencial local apenas ha alcanzado el nivel necesario para generar un potencial de acción, denominado nivel liminar o umbral, se produce sólo después de un período de latencia breve. Un estímulo débil produce un cambio local de potencial en la membrana, aunque la intensidad del potencial local debe aumentar hasta un nivel umbral antes de que se desencadene el potencial de acción.
Período refractario tras un potencial de acción, durante el cual no se puede generar un nuevo estímulo
No se puede producir un nuevo potencial de acción en una fibra excitable mientras la membrana siga despolarizada por el potencial de acción precedente. La única situación que permitirá que se vuelvan a abrir es que el potencial de membrana vuelva al nivel del potencial de membrana en reposo original o cerca del mismo.
El período durante el cual no se puede generar un segundo potencial de acción, incluso con un estímulo intenso, se denomina período refractario absoluto. Para las fibras nerviosas mielinizadas grandes este período es de aproximadamente 1/2.500 s.
Inhibición de la excitabilidad: estabilizadores y anestésicos locales
Los factores estabilizadores de la membrana pueden reducir la exciabilidad. Por ejemplo, una concentración elevada de calcio en el líquido extracelular reduce la permeabilidad de la membrana a los iones sodio y reduce si multáneamente la excitabilidad. Por tanto, se dice que el ion calcio es un estabilizador.
Anestésicos locales
Entre los estabilizadores más importantes están anestésicos locales, como la procaína y tetracaína, que actúa directamente sobre las compuertas de activación de los canales de sodio, haciendo que sea mucho más difícil abrir estas compuertas, reduciendo de esta manera la excitabilidad de la membrana; los impulsos nerviosos no pasan a lo largo de los nervios anestesiados.
Registro de potenciales de membrana y potenciales de acción
Osciloscopio de rayos catódicos
El potencial de membrana cambia muy rápidamente durante el transcurso de un potencial de acción. El único tipo habitual de medidor que puede responder con exactitud a los rápidos cambios del potencial de la membrana es el osciloscopio de rayos catódicos.
El propio tubo de rayos catódicos está formado básicamente por un cañón de electrones y una pantalla fluorescente contra la que se disparan los electrones. Además, el tubo de rayos catódicos está dotado de dos grupos de placas con carga eléctrica, uno situado a ambos lados del haz electrónico y el otro situado por encima y por debajo del mismo. Los circuitos de control electrónico adecuados modifican el voltaje de estas placas, de modo que se puede desviar el haz electrónico hacia arriba o hacia abajo en respuesta a las señales eléctricas que proceden de los electrodos de registro que están situados sobre los nervios. También se puede hacer un barrido horizontal del haz de electrones a través de la pantalla a una frecuencia de tiempo constante por un circuito electrónico interno del
osciloscopio.
El líquido extracelular contiene una gran cantidad de sodio y de iones cloruro, pero poco potasio, lo contrario ocurre en el líquido intracelular, el cual además contiene más fosfatos y proteínas.
La barrera lipídica y las proteínas de transporte de la membrana celular
La membrana celular contiene grandes números de moléculas proteicas insertadas en los lípidos . La bicapa lipídica constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular, aunque hay unas sustancias que pueden penetrarla y difundirse directamente a través de la propia sustancia lipídica. La mayoría de las proteínas penetrantes actúan de formas diferentes. Por ejemplo, las proteínas de los canales y las transportadoras, las primeras permiten el movimiento libre de agua, iones o moléculas a través de la membrana, mientras que las segundas se unen a ellos para transportarlos; sin embargo, ambas son muy selectivas para los tipos de molecúlas o iones que pueden atravesar la membrana.
«Difusión » frente «transporte activo»
El transporte a través de la membrana se produce mediante:
Difusión: la cual se produce por la energía del movimiento cinético normal de la materia.
Transporte activo: las sustancias se combinan con una proteína transportadora para atravesar la menbrana; necesita de otra fuente de energía adicional, además de la cinética.
Difusión
Es un movimiento molecular aleatorio de las sustancias molécula a molécula, a través de espacios intermoleculares de la membrana o en combinación con una proteína transportadora.
Cada partícula se mueve de manera completamente independiente y es lo que los físicos llaman «calor«. Dicho movimiento nunca se interrumpe en ninguna situación salvo a la temperatura de cero absoluto; así pues, las moléculas van adquiriendo y perdiendo energía cinética. El movimiento continuo de moléculas entre sí en los líquidos y en los gases es lo que se llama «difusión«.
Difusión a través de la membrana celular
La difusión a través de la membrana celular se divide en 2 subtipos:
Difusión simple: en ella el movimiento cinético de las moléculas o de los iones se produce a través de una abertura de la membrana o a través de espacios intermoleculares sin ninguna interacción con las proteínas transportadoras de la membrana. Se produce ya sea a través de los intersticios de la bicapa lipídica si la sustancia que difunde es liposoluble o bien por medio de canales acuosos que penetran en todo el grosor de la bicapa a través de las grandes proteínas transportadoras.
Difusión facilitada: es la que necesita la interacción de una proteína transportadora para el paso de las moléculas o iones a través de de la membrana por unión química.
Difusión de sustancias liposoubles a través de la bicapa lipídica
Un factor importante que determina la rapidez con la que una sustancia se difunde a través de la bicapa lipídica es su liposolubilidad. Por ejemplo, el oxígeno, nitrógeno, anhídrido carbónico y los alcoholes tienen una liposolubilidad elevada y va a ser directamente proporcional a la velocidad de difusión de cada uno a través de la membrana.
Difusión de agua y de otras moléculas insolubles en lípidos a través de canales proteicos
El agua pasa rápidamente a través de los canales de las moléculas proteicas que penetran en todo en todo el espesor de la membrana, mientras que otras moléculas insolubles en lípidos pueden atravesar los canales de los poros proteicos de la misma manera que las moléculas de agua si son hidrosolubles y de un tamaño lo suficientemente pequeño, pero a medida que se hacen mayores su penetración disminuye rápidamente.
Difusión a través de poros y canales proteicos: permeabilidad selectiva y activación de canales
Los poros están compuestos por proteínas de membranas celulares integrales que forman tubos abiertos a través de la membrana y que están siempre abiertos. Sin embargo, el diámetro de un poro y sus cargas eléctricas proporcionan una selectividad que permite solo el paso de ciertas moléculas a través de ellos. Un ejemplo claro son las acuaporinas o canales de agua, las cuales solo permiten el paso de agua por las membranas celulares. En las distintas células del cuerpo humano se han descubierto al menos 13 tipos diferentes de acuaporinas. La densidad de algunas acuaporinas en las membranas celulares no es estática, sino que se ve alterada según las diferentes condiciones fisiológicas.
Los canales proteicos se distinguen por su permeabilidad de manera selectiva y por clasificarse en canales activados por voltaje o por ligandos.
Permeabilidad selectiva de los canales proteicos
Mucho de los canales proteicos son muy selectivos sobre todo por su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas.
Los canales de potasio permiten el paso de iones potasio a través de la membrana celular con una facilidad aproximadamente 1.000 veces mayor que para el paso de iones sodio. Se descubrió que los canales de potasio tienen una estructura tetramérica consistente en cuatro subunidades proteicas idénticas que rodean a un poro central. En la parte superior del poro del canal se distribuyen bucles de poro que forman un estrecho filtro de selectividad. Como revestimiento del filtro de selectividad hay oxígenos de carbonilo.
Se cree que existen diferentes filtros de selectividad que determinan la especificidad del canal para cationes o aniones o para iones determinados. Por ejemplo, el canal del sodio mide solo de 0,3 a 0,5 nm de diámetro y sus superficies internas están revestidas con aminoácidos que tienen una carga intensamente negativa, las cuales arrastran pequeños iones de sodio deshidratados hacia el interior de estos canales.
Activación de los canales proteicos
La activación de los canales proteicos proporciona un medio para controlar la permeabilidad iónica de los canales. La apertura y el cierre de las compuertas están controlados de 2 maneras principales:
Activación por voltaje
En ella la conformación molecular de la compuerta o de sus enlaces químicos responde al potencial eléctrico que se establece a través de la membrana celular. Este es el mecanismo básico para generar los potenciales de acción nerviosos que son responsables de las señales nerviosas.
Activación química (por ligando)
En ella las compuertas de algunos canales proteicos se abren por la unión de una sustancia química o ligando a la proteína, que produce un cambio conformacional o un cambio de los enlaces químicos de la molécula de la proteína que abre o cierra la compuerta. Uno de los casos más importantes de activación química es el efecto de la acetilcolina sobre el denominado canal de la acetilcolina, la cual es importante para la transmisión de señales nerviosas de una célula nerviosa a otra o a las células musculares.
Estado abierto frente a estado cerrado de los canales activados
El canal conduce la corriente según un mecanismo de «todo o nada», es decir, la compuerta del canal se abre súbitamente y después se cierra súbitamente, de modo que cada estado abierto dura únicamente desde una fracción de milisegundo hasta varios milisegundos. Esto demuestra la rapidez con la que se producen los cambios durante la apertura y el cierre de las compuertas moleculares proteicas.
Método del pinzamiento zonal de membrana para el registro del flujo de las corrientes iónicas a través de canales aislados
En el método de pinzamiento zonal de membrana se coloca una micropipeta, que tiene un diámetro de sólo 1 o 2 mm, sobre la parte externa de una membrana celular. Después se aplica aspiración en el interior de la pipeta para traccionar la membrana contra la punta de la pipeta. Esto crea un sello en el que los bordes de la pipeta tocan la membrana celular. El resultado es un minúsculo «parche» de membrana en la punta de la pipeta a través del cual se puede registrar el flujo de la corriente eléctrica; alternativamente el pequeño parche de membrana celular y el extremo de la pipeta se pueden separar de la célula. Después se inserta la pipeta con su parche sellado en una solución libre. Esto permite alterar la concentración de los iones tanto en el interior de la micropipeta como en la solución externa, fijándose a voluntad el voltaje entre los dos lados de la membrana, es decir, se puede pinzar a un voltaje dado.
Difusión facillitada
También se denomina difusión mediada por un transportador, en ella la sustancia que se transporta difunde a través de la membrana utilizando una proteína transportadora específica para contribuir al transporte.
En la difusión facilitada la velocidad de difusión se acerca a un máximo, denominado Vmáx, a medida que aumenta la concentración de la sustancia que difunde y está velocidad no puede aumentar por encima de este máximo.
La velocidad a la que se pueden transportar moléculas por este mecanismo nunca puede ser mayor que la velocidad a la que la molécula proteica transportadora puede experimentar el cambio en un sentido y en otro entre sus dos estados, aunque este mecanismo permite que la molécula transportada se mueva en ambas direcciones a través de la membrana.
Entre las sustancias más importantes que atraviesan las membranas celulares mediante difusión facilitada están la glucosa y la mayor parte de los aminoácidos, además es el principal mecanismo mediante el cual la insulina controla la utilización de glucosa por el cuerpo.
Factores que influyen en la velocidad neta de difusión
La velocidad neta de difusión es proporcional a la diferencia de concentración a través de una membrana:la velocidad de difusión neta hacia el interior de la célula es proporcional a la concentración en el exterior menos la concentración en el interior, o:
Difusión neta ∝ (Ce−Ci)
donde Ce es la concentración en el exterior y Ci es la concentración en el interior.
Efecto del potencial eléctrico de membrana sobre la difusión de iones: el «potencial de Nernst»: Si se aplica un potencial eléctrico a través de la membrana las cargas eléctricas de los iones hacen que se muevan a través de la membrana aun cuando no haya ninguna diferencia de concentración que produzca el movimiento. A la temperatura corporal normal (37 °C), la diferencia eléctrica que permitirá que se alcance el equilibrio entre una diferencia de concentración dada de iones univalentes, como los iones de sodio, se puede determinar a partir de la fórmula denominada ecuación de Nernst:
FEM (en milivoltios) = +/-61 log C1/C2
donde:
FEM: es la fuerza electromotriz (voltaje) entre el lado 1 y el lado 2 de la membrana
C1: es la concentración en el lado 1
C2: es la concentración en el lado 2
Efecto de una diferencia de presión a través de la membrana: En ocasiones se produce una gran diferencia de presión entre los dos lados de una membrana permeable. Cuando la presión es mayor en un lado de la membrana que en el otro, significa que la suma de todas las fuerzas de las moléculas que chocan con los canales de ese lado de la membrana es mayor que en el otro lado y la consecuencia es que se dispone de mayores cantidades de energía para producir el movimiento neto de moléculas desde el lado de presión elevada hacia el lado de presión baja.
Ósmosis a través de membranas con permeabilidad selectiva: «difusión neta» de agua
La ósmosis es el movimiento neto del agua que se debe a la producción de una diferencia de la concentración del agua.
La sustancia más abundante que difunde a través de la membrana celular es el agua y normalmente la cantidad que difunde en ambas direcciones está equilibrada de manera tan precisa que se produce un movimiento neto cero de agua, pero en ciertas condiciones se puede producir una diferencia de concentración del agua a través de la membrana, al igual que de otras sustancias. Cuando ocurre esto se produce movimiento neto de agua a través de la membrana celular, haciendo que la célula se hinche o que se contraiga, dependiendo la dirección del movimiento del agua.
La cantidad exacta de presión necesaria para detener la ósmosis se le conoce como presión osmótica.
Importancia del número de partículas osmóticas (concentración molar) en la determinación de la presión osmótica
La presión osmótica que ejercen las partículas de una solución está determinada por el número de partículas por unidad de volumen del líquido; todas las partículas de una solución, independientemente de su masa, ejercen, en promedio, la misma cantidad de presión contra la membrana. El factor que determina la presión osmótica de una solución es la concentración de la solución en función del número de partículas (que es lo mismo que la concentración molar si es una molécula no disociada).
Osmolalidad: el osmol
Un osmol es el peso molecular-gramo de un soluto osmóticamente activo y se utiliza para expresar la concentración de una solución en función del número de partículas en lugar de los gramos.
Una solución que tiene 1 osmol de soluto disuelto por cada kilogramo de agua tiene una osmolalidad de 1 osmol por kilogramo, y una solución que tiene 1/1.000 osmoles disueltos por kilogramo tiene una osmolalidad de 1 mosmol por kilogramo. La osmolaridad normal de los líquidos extracelular e intracelular es de aproximadamente 300 mosmol por kilogramo de agua.
Relación entre osmolalidad y presión osmótica
Muchos de los iones de los líquidos corporales están muy atraídos entre sí, por lo que no se pueden mover totalmente sin restricciones en los líquidos y generar todo su potencial de presión osmótica. Por ello, en promedio la presión osmótica real de los líquidos corporales es de aproximadamente 0,93 veces el valor calculado, el cual es de 5500 mmHg.
El término «osmolaridad»: Es la concentración osmolar expresada en osmoles por litro de solución. Sin embargo, en sentido estricto, es la osmolalidad la que determina la presión osmótica, pero para las soluciones diluidas como las que se encuentran en el cuerpo las diferencias cuantitativas entre la osmolaridad y la osmolalidad son menores del 1%.
Transporte activo de sustancias a través de las membranas
El transporte activo es cuando una membrana celular transporta moléculas o iones «contra corriente» contra un gradiente de concentración o «contra corriente» contra un gradiente eléctrico o de presión.
Entre las sustancias que se transportan activamente a través de algunas membranas celulares están los iones sodio, potasio, calcio, hierro, hidrógeno, cloruro, yoduro y urato, diversos azúcares diferentes y la mayor parte de los aminoácidos.
Transporte activo primario y transporte activo secundario
En el transporte activo primario la energía procede directamente de la división del trifosfato de adenosina (ATP) o de algún otro compuesto de fosfato de alta energía.
En el transporte activo secundario la energía procede secundariamente de la energía que se ha almacenado en forma de diferencias de concentración iónica de sustancias moleculares o iónicas secundarias entre los dos lados de una membrana celular, que se generó originalmente mediante transporte activo primario.
En ambos casos el transporte depende de proteínas transportadoras, las cuales son capaces de impartir energía a la sustancia transportada para moverla contra el gradiente electroquímico.
Transporte activo primario
Entre las sustancias que se transportan mediante transporte activo primario están el sodio, el potasio, el calcio, el hidrógeno, el cloruro y algunos otros iones.
Bomba sodio-potasio
Es el proceso de transporte que bombea iones sodio hacia fuera a través de la membrana celular de todas las células y al mismo tiempo bombea iones potasio desde el exterior hacia el interior.
Es responsable de mantener las diferencias de concentración de sodio y de potasio a través de la membrana celular, así como de establecer un voltaje eléctrico negativo en el interior de las células y es la base de la función nerviosa.
Uno de los componentes físicos de esta bomba es una proteína transportadora, la cual es un complejo formado por dos proteínas globulares distintas: una de mayor tamaño denominada subunidad α y una más pequeña denominada subunidad β. La proteína de mayor tamaño tiene tres características específicas que son importantes para el funcionamiento de la bomba:
Tiene tres puntos receptores para la unión de iones sodio en la porción de la proteína que protruye hacia el interior de la célula.
Tiene dos puntos receptores para iones potasio en el exterior.
La porción interior de esta proteína cerca de los puntos de unión al sodio tiene actividad adenosina trifosfatasa (ATPasa).
Si se aumentan experimentalmente los gradientes electroquímicos de Na+ y de K+ lo suficiente como para que la energía que se almacena en sus gradientes sea mayor que la energía química de la hidrólisis del ATP, estos iones se desplazarán según sus gradientes de concentración y la bomba Na+-K+ sintetizará ATP a partir de ADP y fosfato. Las concentraciones relativas de ATP, ADP y fosfato, así como los gradientes electroquímicos de Na+ y K+, determinan la dirección de la reacción enzimática.
La bomba Na+-K+ es importante para controlar el volumen celular
Una de las funciones más importantes de la bomba Na+-K+ es controlar el volumen de todas las células. Sin la función de esta bomba la mayor parte de las células del cuerpo se hincharía hasta explotar.
La es mucho menos permeable a los iones sodio que a los iones potasio, de modo que una vez que los iones sodio están en el exterior tienen una intensa tendencia a permanecer ahí. Si una célula comienza a hincharse por cualquier motivo, esto automáticamente activa la bomba Na+-K+, moviendo aún más iones hacia el exterior y transportando agua con ellos.
Además se dice que la bomba Na+-K+ es electrógena porque genera un potencial eléctrico a través de la membrana celular.
Transporte activo primario de iones calcio
Los iones calcio normalmente se mantienen a una concentración muy baja en el citosol intracelular de prácticamente todas las células del cuerpo. Esto gracias a dos bombas de calcio que funcionan mediante transporte activo primario. Una de ellas, que está en la membrana celular, bombea calcio hacia el exterior de la célula; la otra bombea iones calcio hacia uno o más de los orgánulos vesiculares intracelulares de la célula. La proteína transportadora penetra en la membrana y actúa como una enzima ATPasa.
Transporte activo primario de iones hidrógeno
El transporte activo primario de los iones hidrógeno es importante en dos localizaciones del cuerpo:
En las glándulas gástricas del estómago, en donde las células parietales que están en las capas profundas tienen el mecanismo activo primario más potente de transporte de iones hidrógeno de todo el cuerpo. Esta es la base para secretar ácido clorhídrico en las secreciones digestivas del estómago.
En la porción distal de los túbulos distales y en los conductos colectores corticales de los riñones, donde hay células intercaladas especiales. Aquí es donde se secretan grandes cantidades de iones hidrógeno desde la sangre hacia la orina con el objetivo de eliminar de los líquidos corporales el exceso de iones hidrógeno.
Energética del transporte activo primario
La cantidad de energía necesaria para transportar activamente una sustancia a través de una membrana viene determinada por cuánto se concentra la sustancia durante el transporte.
Transporte activo secundario: cotransporte y contratransporte
El cotransporte es una forma de transporte activo secundario y se da cuando en condiciones adecuadas la energía de difusión del sodio puede arrastrar otras sustancias junto con el sodio a través de la membrana celular. Para que el sodio arrastre otra sustancia con él es necesario un mecanismo de acoplamiento que se consigue por medio de otra proteína transportadora de la membrana celular. En este caso el transportador actúa como punto de unión tanto para el ion sodio como para la sustancia que se va a cotransportar.
En el contratransporte, los iones sodio intentan una vez más difundir hacia el interior de la célula debido a su gran gradiente de concentración. Sin embargo, esta vez la sustancia que se va a transportar está en el interior de la célula y se debe transportar hacia el exterior.
Cotransporte de glucosa y aminoácidos junto con iones sodio
La glucosa y muchos aminoácidos se transportan hacia el interior de la mayor parte de las células contra grandes gradientes de concentración. La proteína transportadora tiene dos puntos de unión en su cara externa, uno para el sodio y otro para la glucosa. Cuando ambos están unidos se produce automáticamente el cambio conformacional y el sodio y la glucosa son transportados al mismo tiempo hacia el interior de la célula. Por tanto, este es un mecanismo de cotransporte sodio-glucosa.
El cotransporte con sodio de los aminoácidos se produce de la misma manera que para la glucosa, excepto porque utiliza un grupo diferente de proteínas transportadoras. Se han identificado cinco proteínas transportadoras de aminoácidos.
El cotransporte con sodio de la glucosa y de los aminoácidos se produce especialmente a través de las células epiteliales del tubo digestivo y de los túbulos renales para favorecer la absorción de estas sustancias hacia la sangre.
Contratransporte con sodio de iones calcio e hidrógen
Dos mecanismos de contratransporte especialmente importantes son el contratransporte sodio-calcio, el cual se produce a través de todas o casi todas las membranas celulares, y el contratransporte sodio-hidrógeno que se produce en varios tejidos. Un ejemplo especialmente importante se produce en los túbulos proximales de los riñones.
El contratransporte puede transportar cantidades muy grandes de iones hidrógeno, lo que hace que sea clave para el control del ion hidrógeno en los líquidos corporales.
Transporte activo a través de capas celulares
El transporte de este tipo se produce a través de:
El epitelio intestinal
El epitelio de los túbulos renales
El epitelio de todas las glándulas exocrinas
El epitelio de la vesícula biliar
La membrana del plexo coroideo del cerebro y otras membranas
El mecanismo básico para el transporte de una sustancia a través de una lámina celular es:
Transporte activo a través de la membrana celular de un polo de las células transportadoras de la capa.
Difusión simple o difusión facilitada a través de la membrana del polo opuesto de la célula.
El transporte activo de los iones sodio en las superficies basolaterales de las células epiteliales da lugar a transporte no sólo de iones sodio, sino también de agua. Estos son los mecanismos mediante los cuales casi todos los nutrientes, los iones y otras sustancias se absorben hacia la sangre desde el intestino; también son la forma en la que algunas sustancias se reabsorben desde el filtrado glomerular por los túbulos renales.
Los genes, situados en el núcleo de todas las células del organismo, controlan la herencia de padres a hijos, pero también son los encargados de controlar las funciones de la célula determinando qué sustancias se sintetizan dentro de ésta, es decir, qué estructuras, qué enzimas y qué productos químicos participan.
Cada gen compuesto por ácido desoxirribonucleico (ADN) controla la formación del ácido ribonucleico (ARN), mismo que se esparcirá en la celúla para poder formar una proteína. El proceso completo, desde la transcripción del código genético en el núcleo hasta la traducción del código del ARN y la formación de proteínas en el citoplasma celular se conoce como expresión génica.
Hay cerca de 30000 genes diferentes por cada célula, lo que hace posible formar un total de casi 100000 diversos tipos de proteínas celulares. Algunas de ellas son proteínas estructurales, aunque la mayoría son enzimas que catalizan las reacciones químicas de la célula.
Genes en el núcleo celular
Bloques básicos de ADN
Los compuestos químicos básicos que forman el el ADN son:
El ácido fosfórico
El azúcar desoxirribosa
Cuatro bases nitrogenadas: dos purínicas (adenina y guanina) y dos pirimidínicas, (timina y citosina)
El ácido fosfórico y la desoxirribosa forman las dos hebras helicoidales que sirven de soporte para la molécula de ADN, mientras que las bases nitrogenadas se apoyan entre las dos hebras y se conectan entre sí.
Nucleótidos
La primera etapa en la formación del ADN consiste en combinar una molécula de ácido fosfórico, una molécula de desoxirribosa y una de las cuatro bases para formar un nucleótido ácido, creando uno por cada base:
Desoxiadenílico
Desoxitimidílico
Desoxiguanílico
Desoxicitidílico
Los nucleótidos se organizan para formar dos hebras de ADN unidas laxamente entre sí
El esqueleto de cada hebra de ADN está compuesto por moléculas de ácido fosfórico y desoxirribosa que se van alternando, mientras las bases de purina y pirimidina se unen a los lados de las moléculas de desoxirribosa. Después, las dos hebras respectivas de ADN se mantienen unidas mediante enlaces débiles de hidrógeno entre las bases purínicas y pirimidínicas, debido a la laxitud de estos enlaces las dos hebras se separan con facilidad sobre todo cuando realizan sus funciones en la célula. Cabe destacar que:
Cada base purínica de adenina de una hebra siempre se une con una base pirimidínica de timina de la otra.
Cada base purínica de guanina siempre se une con una base pirimidínica de citosina.
En cada vuelta completa de la hélice de la molécula de ADN hay 10 pares de nucleótidos.
Código genético
La importancia del ADN se debe a su capacidad para controlar la formación de las proteínas en la célula mediante un código genético, mismo que consta de tripletessucesivos de bases, es decir, tres bases sucesivas componen una palabra del código. Los tripletes sucesivos controlan en último término la secuencia de aminoácidos en una molécula proteica que la célula debe sintetizar.
El código de ADN del núcleo celular se transfiere al código de ARN en el citoplasma celular: proceso de transcripción
Para que los genes de ADN del núcleo controlen las reacciones químicas del citoplasma se necesita de la intervención del ARN. El código se transfiere al ARN en un proceso que se conoce como transcripción y se difunde desde el núcleo a través de los poros del mismo al compartimiento citoplasmático, donde controla la síntesis proteica.
Síntesis de ARN
Durante la síntesis de ARN las dos hebras de la molécula de ADN se separan temporalmente y una de ellas se usa como plantilla para la síntesis de una molécula de ARN. Los tripletes del código del ADN provocan la formación de tripletes con un código complementario (o codones) en el ARN; a su vez, estos codones controlarán la secuencia de aminoácidos en una proteína que se va a sintetizar en el citoplasma celular.
Bloques básicos para la construcción del ARN
En la formación del ARN no se usa el azúcar desoxirribosa, en cambio se utiliza la ribosa, que contiene un ion hidroxilo extra unido a la estructura anular de la ribosa. Además, la timina se reemplaza por otra pirimidina, el uracilo.
Formación de nucleótidos de ARN
Los bloques básicos de ADN forman los nucleótidos de ARN. Se usan también cuatro nucleótidos distintos para formar el ARN, nucleótidos que contienen las bases adenina, guanina, citosina y uracilo, éste último reemplazando a la timina del ADN.
Activación de los nucleótidos de ARN
El siguiente paso de la síntesis de ARN es la «activación» de los nucleótidos de ARN por una enzima, polimerasa de ARN, añadiendo a cada nucleótido dos radicales fosfato más para formar trifosfatos. Estos dos últimos fosfatos se combinan con el nucleótido mediante enlaces de fosfato de alta energía derivados del ATP celular. Esto hace que cada uno de los nucleótidos puede disponer de grandes cantidades de energía del ATP para favorecer las reacciones químicas que van añadiendo cada nuevo nucleótido de ARN al extremo de la cadena de ARN que se está desarrollando.
Montaje de la cadena de ARN a partir de los nucleótidos activados usando una cadena de ADN como plantilla: proceso de TRANSCRIPCIÓN
La polimerasa de ARN es una gran enzima proteica que tiene muchas propiedades funcionales para la formación de la molécula de ARN, como son:
En la cadena de ADN hay una secuencia de nucleótidos que se denomina promotor. La polimerasa de ARN reconoce este promotor y se une a él en un paso esencial para iniciar la formación de la molécula de ARN.
La polimerasa de ARN, después de unirse al promotor, provoca el desenrollamiento de dos vueltas de la hélice de ADN y la separación de las porciones abiertas de las dos hebras.
La polimerasa se desplaza a lo largo de la hebra de ADN, desenrollando y separando temporalmente las dos hebras de ADN en cada etapa de su movimiento. A medida que se desplaza va añadiéndose en cada paso un nucleótido de ARN activado nuevo en el extremo de la cadena de ARN que se va formando.
El código presente en la cadena de ADN se transmite finalmente de forma complementaria a la cadena de ARN. Las bases de nucleótido con ribosa siempre se combinan con las bases con desoxirribosa, de la siguiente forma:
Tipos de ARN
ARN mensajero precursor (pre-ARNm), que es un gran ARN de cadena única inmaduro que se procesa en el núcleo para formar ARN mensajero (ARNm) maduro. El pre-ARN incluye dos tipos diferentes de segmentos denominados intrones, que son eliminados por un proceso de corte y empalme (splicing), y exones, que se conservan en el ARNm final.
ARN nuclear pequeño (ARNnp), que dirige el corte y empalme de pre-ARNm para formar ARNm.
ARN mensajero (ARNm), que transporta el código genético al citoplasma para controlar el tipo de proteína que se forma.
ARN de transferencia (ARNt), que transporta los aminoácidos activados a los ribosomas para usarlos en el montaje de la molécula proteica.
ARN ribosómico, que, junto con 75 proteínas distintas, forma ribosomas, las estructuras físicas y químicas en las que se montan realmente las moléculas proteicas.
MicroARN (ARNmi), que son moléculas de ARN monocatenario de 21 a 23 nucleótidos capaces de regular la transcripción y la traducción génicas.
ARN mensajero: los codones
Las moléculas de ARNm son cadenas largas y sencillas que se encuentran en suspensión en el citoplasma. Estas moléculas están compuestas por varios cientos a miles de nucleótidos de ARN en cadenas no pareadas y contienen codones que son exactamente complementarios a los tripletes del código de los genes de ADN.
Codones de ARN para los distintos aminoácidos
Un codón representa la señal de iniciar la fabricación de la molécula proteica y tres codones representan la señal de detener la fabricación de la molécula proteica.
ARN de transferencia: los anticodones
El ARNt transfiere las moléculas de aminoácidos a las moléculas proteicas a medida que se va sintetizando la proteína. Cada tipo de ARNt se combina específicamente con 1 de los 20 aminoácidos que se van a incorporar en las proteínas; actúa como vehículo para transportar su tipo específico de aminoácido a los ribosomas, en donde cada tipo específico de ARN de transferencia reconoce un codón en particular en el ARNm y libera el aminoácido apropiado en el lugar apropiado de la cadena de la molécula proteica que se está formando. El ARNt contiene sólo 80 nucleótidos. En uno de los extremos de la molécula siempre hay un ácido adenílico, en cuyo grupo hidroxilo de la ribosa del ácido adenílico se une el aminoácido transportado.
Es esencial que cada tipo de ARNt también sea específico de un codón en particular del ARNm. El código específico del ARNt que le permite reconocer un codón específico es, de nuevo, un triplete de bases de nucleótidos que se denomina anticodón y se sitúa aproximadamente en la zona media de la molécula del ARNt.
Durante la formación de la molécula proteica las bases del anticodón se combinan laxamente mediante enlaces hidrógeno con las bases del codón del ARNm.
ARN ribosómico
Constituye el 60% del ribosoma, el cual es la estructura física del citoplasma en el que se sintetizan realmente las moléculas proteicas; sin embargo, siempre funciona asociado a otros tipos de ARN, es decir, actúa como una planta de fabricación en la cual se forman las moléculas proteicas.
Formación de ribosomas en el nucléolo
Los genes de ADN que se utilizan para la formación del ARN ribosómico se sitúan en cinco pares de cromosomas del núcleo y cada uno de estos contiene muchos duplicados de estos genes en particular. A medida que se forma el ARN ribosómico se recoge en el nucléolo.
El ARN ribosómico se procesa especialmente en el nucléolo donde se une a las proteínas ribosómicas para formar productos de condensación granulares que son las subunidades primordiales de los ribosomas. Estas subunidades se liberan desde el nucléolo y se transportan hasta prácticamente todas las partes del citoplasma. Cuando entran en el citoplasma se reúnen para formar los ribosomas maduros y funcionantes. Por tanto, las proteínas se forman en el citoplasma de la célula, pero no en el núcleo celular, porque el núcleo no contiene los ribosomas maduros.
MicroARN
Se trata de cortos fragmentos de ARN monocatenario (de 21 a 23 nucleótidos) que regulan la expresión génica; se codifican a partir del ADN transcrito de genes, pero no se traducen a proteínas y también reciben el nombre de ARN no codificante.
La generación de ARNmi supone un procesamiento especial de ARN precursores primarios más largos llamados ARNmipri, que son los transcriptos primarios del gen. Los ARNmi-pri se procesan en el núcleo celular por parte del complejo de microprocesador en pre-ARNmi. Estos son procesados después adicionalmente en el citoplasma por una enzima dicer específica que ayuda a ensamblar un complejo de silenciamiento inducido por ARN (RISC) y genera ARNmi.
Los ARNmi desempeñan un papel importante en la regulación normal de la función celular, y las alteraciones en la función de los ARNmi se han asociado con enfermedades como el cáncer y las cardiopatías.
Otro tipo de microARN es el ARN de interferencia pequeño (ARNsi), también denominado ARN de silenciamiento o ARN de interferencia corto. Son moléculas cortas de ARN bicatenario, de 20 a 25 nucleótidos de longitud, que interfieren con la expresión de genes específicos. Los ARNsi se refieren en general a ARNmi sintéticos y pueden administrarse para silenciar la expresión de genes específicos; además, es posible usarlos para bloquear la traducción de cualquier ARNm y, por tanto, la expresión por cualquier gen para el cual se conozca la secuencia de nucleótidos.
Formación de proteínas en los ribosomas: el proceso de TRADUCCIÓN
Cuando una molécula de ARN mensajero entra en contacto con un ribosoma, se desplaza por el mismo a partir de un extremo predeterminado de la molécula de ARN que se
especifica mediante el codón iniciador de la cadena. Mientras el ARN mensajero se desplaza por el ribosoma, se forma una molécula proteica, en un proceso que se conoce como traducción. Después, se marca el final de una molécula proteica cuando el codón terminador de la cadena atraviesa el ribosoma y la molécula proteica se libera en el citoplasma.
Polirribosomas
Una molécula sencilla de ARN mensajero puede formar moléculas proteicas en varios ribosomas al mismo tiempo. Las moléculas proteicas se encuentran en distintas fases de desarrollo en cada ribosoma. Como consecuencia de este proceso es frecuente encontrar agrupaciones de ribosomas, uniéndose entre 3 y 10 ribosomas a una única molécula de ARN mensajero al mismo tiempo. Estos grupos se conocen como polirribosomas.
El ARN mensajero da lugar a la formación de una molécula proteica en cualquier ribosoma, es decir, no hay una especificidad de los ribosomas por un tipo dado de proteína.
Muchos ribosomas se unen al retículo endoplásmico
Los extremos iniciales de muchas moléculas proteicas en formación tienen secuencias de aminoácidos que se unen inmediatamente a los locus de receptores específicos en el retículo endoplásmico; penetran en la pared del retículo hasta la matriz dando un aspecto granular a estas porciones del retículo en las que se están formando las proteínas y entran en la matriz.
Sin embargo, la mayoría de las proteínas que se sintetizan en los ribosomas, excepto en las células glandulares, se liberan directamente al citosol en lugar de al retículo endoplásmico. Estas proteínas son enzimas y proteínas estructurales internas de la célula.
Pasos químicos de la síntesis proteica
Cabe mencionar que se usan cuatro enlaces de alta energía en total por cada aminoácido que se añade a la cadena de la proteína, es decir, la síntesis proteica es uno de los procesos que consume más energía en la célula.
Enlace peptídico
Los aminoácidos sucesivos de la cadena de proteína se combinan entre sí según una reacción típica:
En esta reacción se elimina un radical hidroxilo (OH–) de la porción COOH del primer aminoácido y se elimina un hidrógeno (H+) de la porción NH2 del otro aminoácido. Ambos radicales se combinan para formar agua y los dos locus reactivos que quedan en los dos aminoácidos sucesivos se unen entre sí, con lo que se obtiene una molécula única. Este proceso se conoce como enlace peptídico.
Control de la función génica y actividad bioquímica de las células
Los genes controlan tanto las funciones físicas como las químicas de las células, aunque también debe controlarse el grado de activación de los genes respectivos. Cada célula utiliza mecanismos internos de retroalimentación muy potentes que mantienen el orden de las distintas actividades celulares. Hay dos métodos de control de las actividades químicas de la célula:
REGULACIÓN GENÉTICA
REGULACIÓN ENZIMÁTICA
Regylación genética
Cubre todo el proceso, desde la transcripción del código genético en el núcleo hasta la formación de proteínas en el citoplasma. La regulación diferencial de la expresión génica permite también que los numerosos tipos distintos de células del cuerpo realicen cada uno funciones especializadas.
La medida definitiva de la expresión génica es si se forman productos génicos (proteínas) y en qué medida. Puede tener lugar en cualquier punto de las rutas de transcripción, procesamiento de ARN y traducción.
El promotor controla la expresión génica
La transcripción de ADN está controlada por elementos reguladores encontrados en el promotor de un gen. En eucariotas el promotor basal consta de una secuencia de siete bases (TATAAAA) denominada caja TATA, sitio de unión para la proteína de unión a TATA, y otros varios e importantes factores de transcripción que se conocen conjuntamente como complejo IID del factor de transcripción. . Este promotor basal está presente en todos los genes de codificación de proteínas y la polimerasa debe unirse con este antes de que pueda empezar a desplazarse a lo largo de la cadena del ADN para sintetizar el ARN. El promotor corriente arriba está situado antes del lugar de inicio de la transcripción y contiene varios sitios de unión para factores de transcripción positivos y negativos.
En la transcripción de genes en eucariotas influyen también los reforzadores, que son regiones de ADN que pueden unirse a factores de transcripción y aunque pueden estar ubicados a una gran distancia de su gen objeto, cuando el ADN se encuentra arrollado en el núcleo pueden hallarse relativamente cerca. Se estima que hay 110.000 secuencias de reforzadores génicos en el genoma humano.
Por otro lado, también hay aisladores cromosómicos, los cuales son secuencias génicas que proporcionan una barrera de tal forma que un gen específico queda aislado de las influencias de transcripción de los genes circundantes. Los aisladores son enormemente variables en su secuencia de ADN y en las proteínas que se unen a ellos. Una forma posible de modulación de la actividad de un aislador es la metilación de ADN.
Los sistemas de control de los genes son especialmente importantes para controlar las concentraciones intracelulares de aminoácidos.
Control de las funciones intracelulares mediante la regulación enzimática
La regulación enzimática representa una segunda categoría de mecanismos por los cuales se pueden controlar las funciones bioquímicas celulares.
Inhibición enzimática
Algunas de las sustancias químicas formadas en la célula ejercen una retroalimenta ción directa inhibiendo los sistemas enzimáticos específicos que los sintetizan. Casi siempre, el producto sintetizado actúa sobre la primera enzima de una secuencia en lugar de sobre las enzimas sucesivas, impidiendo la acumulación de los productos intermedios que no se están usando. La inhibición enzimática es otro ejemplo de control mediante retroalimentación negativa.
Activación enzimática
Las enzimas que están normalmente inactivas se activan cuando es necesario.
El AMPc actúa como un activador enzimático de la enzima fosforilasa y, por tanto, facilita el control de la concentración intracelular de ATP.
Las purinas formadas inhiben las enzimas que se requieren para la formación de nuevas purinas, aunque activan las enzimas necesarias para la formación de pirimidinas. Por el contrario, las pirimidinas inhiben sus propias enzimas, pero activan las enzimas de las purinas.
El sistema genético de ADN también controla la reproducción celular
El sistema genético controla cada etapa del desarrollo del ser humano; si hay un eje central a la vida, es el sistema genético del ADN.
Ciclo vital de la célula
El ciclo vital de una célula es el período que transcurre desde el inicio de la reproducción celular hasta el inicio de la siguiente. En las células de los mamíferos su ciclo vital puede ser de 10 a 30 h y termina por una serie de sucesos físicos diferenciados que se denominan mitosis y que tienen como finalidad lograr la división de la célula en dos células hijas nuevas. La mitosis en sí misma dura sólo unos 30 min, por lo que más del 95% del ciclo vital de las células está representado por el intervalo entre las mitosis, o interfase, incluso en las células que se reproducen con mayor rapidez. Los factores inhibidores casi siempre disminuyen la velocidad o detienen el ciclo vital no inhibido.
La reproducción celular comienza con la replicación del ADN
La reproducción comienza en el núcleo. El primer paso consiste en la replicación de todo el ADN de los cromosomas y sólo después puede tener lugar la mitosis, el ADN comienza a duplicarse entre 5 y 10 h antes de ésta y se completa en 4-8 h. Después de esta replicación hay otro periodo de 1-2 h antes de que comience bruscamente la mitosis.
Reparación de ADN, corrección de lectura y mutaciones del ADN
Durante la hora que transcurre entre la replicación del ADN y el comienzo de la mitosis hay un período de reparación muy activa y «corrección de lectura» de las cadenas de ADN. Para ello se utilizan las mismas polimerasas del ADN y ADN ligasas que se usan en la replicación. Este proceso de reparación del ADN se conoce como corrección de lectura.
No obstante, el proceso de transcripción comete errores pocas veces y cuando lo hace el error se denomina mutación, misma que provoca la formación de alguna proteína anormal en la célula en lugar de la proteína necesaria, lo que conduce a funciones celulares anormales y, en ocasiones, la muerte celular. Por ello, como protección el genoma humano está representado por dos conjuntos independientes de cromosomas con genes casi idénticos.
Cromosomas y su replicación
Las hélices de ADN del núcleo se enrollan en cromosomas. La célula humana contiene 46 cromosomas dispuestos en 23 pares. Los genes de los dos cromosomas de cada par son idénticos o casi idénticos entre sí en su mayor parte. Tienen una gran cantidad de proteínas, principalmente muchas moléculas pequeñas de histonas cargadas positivamente, estas se organizan en un número inmenso de pequeños núcleos alrededor de los cuales se enrollan secuencialmente pequeños segmentos de cada hélice de ADN. Se ha demostrado que algunas de las proteínas reguladoras son capaces de disminuir la densidad del enrollamiento del ADN en las histonas y permitir que pequeños segmentos cada vez permitan la formación del ARN.
La replicación de todos los cromosomas se produce en los minutos siguientes a la finalización de la replicación de las hélices de ADN. Los dos cromosomas recién formados se mantienen unidos entre sí (hasta el momento de la mitosis) en un punto que se denomina centrómero, situado cerca del centro. Estos cromosomas duplicados, pero aún unidos entre sí, se conocen como cromátidas.
Mitosis celular
Es el proceso por el que la célula se divide en dos células nuevas. Se produce automáticamente en 1 o 2 horas.
Aparato mitótico: función de los centriolos
Uno de los primeros pasos de la mitosis tiene lugar en el citoplasma en torno o en unas pequeñas estructuras denominadas centríolos. Cada centríolo es un pequeño organismo cilíndrico en torno a 0,4 mm de largo y 0,15 mm de diámetro y está formado principalmente por nueve estructuras tubulares paralelas dispuestas en forma de un cilindro.
Dos pares de centríolos se mantienen estrechamente unidos cerca de un polo del núcleo y cada par de centríolos, junto al material pericentriolar unido a él, compone el centrosoma.
Poco antes de la mitosis, los dos pares de centriolos comienzan a separarse uno de otro, a su vez que crecen radialmente otros microtúbulos que alejan los pares de centríolos, formando una estrella a modo de soporte, que se conoce como áster, en cada extremo de la célula.
El complejo de microtúbulos que se extiende entre los dos nuevos pares de centríolos es el huso, y todo el conjunto de microtúbulos más los dos pares de centríolos se denomina aparato mitótico.
Etapas de la mitosis
Profase
Prometafase
Metafase
Anafase
Telofase
Control del crecimiento y la reproducción celular
Algunas células crecen y se reproducen continuamente, mientras que otras pueden no reproducirse durante muchos años, como las neuronas y la mayoría de los miocitos de del músculo estriado.
En los estudios experimentales se ha demostrado al menos tres formas de controlar el crecimiento:
El crecimiento se controla a menudo me diante factores de crecimiento que proceden de otras partes del organismo.
La mayoría de las células normales dejan de crecer cuando han salido de este espacio para su crecimiento, lo que sucede cuando las células crecen en un cultivo tisular.
Las células que crecen en un cultivo celular dejan de crecer cuando se ha podido re coger cantidades diminutas de sus propias secreciones en el medio de cultivo, lo cual, además, sería un medio de controlar el crecimiento mediante la retroalimentación negativa.
Regulación del tamaño de la célula
El tamaño de la célula está determinado casi en su totalidad por la cantidad de ADN funcionante que hay en el núcleo.
Es posible que, usando el quimioterápico colchicina, se impida la formación del huso mitótico y, por tanto, la mitosis aunque continúe la replicación del ADN. En este caso, el núcleo contiene una cantidad de ADN bastante mayor de lo que normalmente contendría y la célula crece hasta un tamaño proporcionalmente mayor.
Diferenciación celular
La diferenciación celular se refiere a los cambios de las propiedades físicas y funcionales de las células a medida que proliferan en el embrión para formar las distintas estructuras y órganos corporales; es consecuencia no de la pérdida de genes, sino de la represión selectiva de los distintos promotores génicos.
El genoma celular comienza a producir en una cierta etapa de la diferenciación celular una proteína reguladora que reprimirá para siempre a un grupo selecto de genes, es decir, los genes reprimidos no volverán a funcionar. Las células humanas maduras producen un máximo de 8.000 a 10.000 proteínas .
Los estudios embriológicos demuestran que algunas células del embrión controlan la diferenciación de las células adyacentes.
Apoptosis: muerte celuñar programada
Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis.
Es una muerte celular metódica que da lugar al desmontaje y fagocitosis de la célula antes de que se produzca ninguna fuga de su contenido, por lo que las células vecinas se mantienen sanas. Se inicia mediante la activación de una familia de proteasas que se conocen como caspasas. Se trata de unas enzimas que se sintetizan y almacenan en la célula en forma de procaspasas inactivas.
Por otro lado, las células que mueren como consecuencia de una lesión aguda se hinchan y estallan debido a la pérdida de la integridad de la membrana celular, un proceso que se denomina necrosis celular.
En los seres humanos adultos miles de millones de células mueren cada hora en tejidos como el intestino y la médula ósea y se reemplazan con células nuevas. No obstante, en los adultos sanos la muerte celular programada está normalmente equilibrada con la formación de células nuevas, ya que, de lo contrario, los tejidos del organismo aumentarían o disminuirían excesivamente.
La apoptosis pueden ser importantes en las enfermedades neurodegenerativas, por lo que algunos fármacos que se han usado con éxito para la quimioterapia inducen apoptosis en las células cancerosas.
Cáncer
Se debe en todos o casi todos los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los genes anormales se denominan oncogenesy se han descu bierto hasta 100 tipos distintos., aunque también hay antioncogenes, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer.
La probabilidad de que alguien desarrolle un cáncer se multiplica hasta por cinco en personas en las que se suprimen los sistemas inmunitarios.
Además, la probabilidad de mutaciones aumenta muchas veces cuando una persona se expone a determinados factores químicos, físicos o biológicos, como son los siguientes:
La radiación ionizante, como los rayos X, los rayos gamma y la radiación de partículas procedentes de sustancias radiactivas, e incluso la luz ultravioleta.
Las sustancias químicas que provocan la mutación se denominan carcinógenos. Los carcinógenos que actualmente provocan el mayor número de muertes son los contenidos en el humo de los cigarrillos.
Los irritantes físicos, pues el daño de los tejidos conduce a una sustitución mitótica rápida de las células.
Tendencia hereditaria, lo que es consecuencia de que la mayoría de los cánceres requieren no sólo una mutación, sino dos o más antes de que aparezca el cáncer.
Algunos virus provocan ciertas clases de cáncer, como la leucemia.
Características invasivas de la célula cancerosa
La célula cancerosa no respeta los límites habituales del crecimiento celular.
Las células cancerosas son bastante menos adhesivas entre sí que las células normales, por lo que tienden a dispersarse por los tejidos, entrar al torrente sanguíneo y transportarse por el organismo.
Algunos cánceres también producen factores angiogénicos que provocan el crecimiento de muchos vasos sanguíneos nuevos dentro del cáncer.
¿Por qué matan las células cancerosa?
El tejido canceroso compite con los tejidos normales para lograr los nutrientes y como continúan proliferando indefinidamente demandan prácticamente toda la nutrición disponible para el organismo o para una parte esencial del mismo. En consecuencia, los tejidos normales sufren una muerte nutritiva gradual.
«Organización funcional del cuerpo humano y control del medio interno»
La fisiología es la ciencia encargada de describir tanto los procesos físicos y químicos que dan origen y desarrollo a la vida. Existe una división para las funciones fisiológicas dependiendo del tipo de vida del que estemos tratando y de las características funcionales que éstas tengan, así por ejemplo se puede hablar de fisiología celular, vegetal, bacteriana, humana, entre otras.
Fisiología humana
Nosotros somos seres vivos por el simple hecho de que podemos percibir, sentir y aprender de las diversas situaciones que se nos presentan, es lo que nos permite existir y hacen posible la vida y justamente la ciencia que trata de estudiar los mecanismos y características del cuerpo humano que lo definen como «ser vivo» es la fisiología humana.
Las células como unidades vivas del cuerpo
La unidad viva básica del cuerpo es la célula. Cada órgano es un agregado de muchas células diferentes que se mantienen unidas mediante estructuras de soporte intercelulares.
El cuerpo humano contiene cerca de 100 billones de células, de las cuales los eritrocitos son una de las más abundantes y a pesar de que cada una de estas células son diferentes entre sí tienen ciertas carecterísticas en común. Por ejemplo, poseen prácticamente los mismos mecanismos químicos generales que permiten cambiar los nutrientes en energía y liberan los productos de sus reacciones químicas en los líquidos circundantes. Además, casi todas tienen la capacidad de reproducirse formando más células de su propia familia, regenerándose cuando éstas se van destruyendo.
Líquido extracelular e intracelular
El cuerpo humano del adulto es 60% líquido, del cual la mayoría es líquido intracelular, mismo que contiene grandes cantidades de iones potasio, magnesio y fosfato. Sin embargo, casi una tercera parte está fuera de las células, es decir, es liquído extracelular o también conocido como «medio interno del organismo», estando en constante movimientoa lo largo de todo el cuerpo. En éste se encuentran los iones y nutrientes que necesitan las células para mantenerse vivas, por ejemplo: oxígeno, glucosa, aminoácidos, sustancias grasas y otros componentes.
Mecanismos homeostáticos de los principales sistemas funcionales
Homeostasis
Grandes fisiólogos, como Walter Cannon, definen homeostasis como el mantenimiento de unas condiciones casi constantes del medio interno. Para que dichas condiciones sean constantes tanto órganos comos tejidos realizan en conjunto diferentes funciones. Este equilibrio es necesario para el mantenimiento de la vida.
Cuando se rompe la homeostasis se dice que hay enfermedad, sin embargo aún en esta situación los mecanismos homeostáticos siguen activos. La fisiopatología pretende explicar cómo se alteran los diversos procesos fisiológicos durante las enfermedades y las lesiones.
Transporte en el líquido extracelular y sistema de mezcla: el aparato circulatorio
Existen dos etapas por las cuales se transporta el líquido extracelular en el organismo:
Movimiento de la sangre por el cuerpo dentro de los vasos sanguíneos
Movimiento del líquido entre los capilares sanguíneos y los espacios intercelulares entre las células tisulares
Conforme la sangre atraviesa los capilares sanguíneos se produce también un intercambio continuo de líquido extracelular entre la porción del plasma de la sangre y el líquido intersticial que rellena los espacios intercelulares. Las paredes de los capilares son permeables casi a todo, excepto por las moléculas proteínas plasmáticas. El líquido se difunde gracias al movimiento cinético de las moléculas en el plasma y en el
líquido intersticial, además de que se va mezclando periodicamente para mantener su homogeneidad.
Origen de los nutrientes en el líquido extracelular
Aparato respiratorio: la sangre pasa por los pulmones y capta el oxígeno a través de los alvéolos, adquiriendo el oxígeno que necesitan las células. La membrana alveolar tiene un grosor de 0,4 a 2 mm y el oxígeno difunde rápidamente por el movimiento molecular a través de esta membrana para entrar en la sangre.
Aparato digestivo: la sangre atraviesa las paredes del aparato digestivo, absorbiendo los distintos nutrientes del alimento al líquido extracelular de la sangre.
Hígado y otros órganos que realizan principalmente funciones metabólicas: el hígado hace utilizables muchas de las sustancias absorbidas cambiando su composición química, mientras que otros tejidos las almacenan, modifican o eliminan.
Aparato locomotor: el organismo se puede muever en espacio y tiempo adecuado gracias a los músculos, los cuales también nos brinda protección para el enterno.
Eliminación de los productos finales metabólicos
Eliminación del dióxido de carbono en los pulmones: el dióxido de carbono es el producto final más abundante del metabolismo y se libera de la sangre a los álveolos, los pulmones y, por último, hacia la atmósfera.
Los riñones: Tienen la función de filtrar, reabsorber o eliminar sustancias del cuerpo, evitando su acumulación en el líquido extracelular.
Aparato digestivo: El material no digerido que entra en el aparato digestivo y algunos productos residuales del metabolismo se eliminan en las heces.
Hígado: Ayuda a detoxificar o eliminar fármacos y productos químicos que se ingieren. Secretando sus residuos en la bilis para su eliminación en las heces.
Regulación de las funciones corporales
Sistema nervioso: compuesto por 3 partes:
Porción de aferencia sensitiva
El sistema nervioso central (o la porción integradora)
La porción eferente motora
El sistema nervioso central está formado por el cerebro y la médula espinal. También está el sistema nervioso autónomo o neurovegetativo que funciona subconscientemente y controla funciones de los órganos internos.
Sistemas hormonales: Regula muchas funciones metabólicas a través de las hormonas, proporcionando un sistema de regulación complementario al sistema nervioso. Existen 8 glándulas endocrinas mayores.
Protección del cuerpo
Sistema inmunitario: proporciona un mecanismo para que el cuerpo diferencie sus propias células de las extrañas y destruya al invasor por fagocitosis o mediante la producción anticuerpos, destruyendo o neutralizando al invasor.
Sistema tegumentario: cubre, amortigua, proteje a tejidos y órganos, regula la temperatura y la excreción de sustancias, delimita el medio corporal interno del mundo exterior.
Reproducción
Mantiene la homeostasis al crear nuevos seres que sustituirán a los que mueren, ayudando a mantener el automatismo y la continuidad de la vida.
Sistemas de control del organismo
El cuerpo humano contiene miles de sistemas de control, algunos actúan dentro de los órganos para controlar las funciones de cada componente de los mismos, otros actúan a través de todo el organismo para controlar las interrelaciones entre los órganos.
Ejemplos de mecanismos de control
Regulación de las concentraciones de oxígeno y dióxido de carbono en el líquido extracelular: el organismo cuenta con un mecanismo de control especial para mantener una concentración constante de oxígeno en el líquido extracelular, mismo que depende principalmente de las características químicas de la hemoglobina. La regulación de la concentración de oxígeno en los tejidos se basa en las características químicas de la propia hemoglobina, regulación que se conoce como función amortiguadora de oxígeno de la hemoglobina. Por otro lado, el dióxido de carbono es el principal producto final de las reacciones oxidativas de las células, una concentración mayor de lo normal de dióxido de carbono en la sangre excita el centro respiratorio, lo que aumenta la eliminación del exceso de éste en la sangre y los líquidos tisulares.
Regulación de la presión arterial:
Sistema de barorreceptores.- Cuando la presión arterial es demasiado elevada los barorreceptores envían descargas de impulsos nerviosos al bulbo raquídeo cerebral, mientras que al mismo tiempo disminuyen el número de impulsos transmitidos desde el centro vasomotor a través del sistema nervioso simpático hacia el corazón y los vasos sanguíneos. La ausencia de estos hace que disminuya la actividad de bomba en el corazón y produce una dilatación de los vasos sanguíneos periféricos, aumentando el flujo de sangre a través de ellos. Ambos efectos hacen que la presión arterial disminuya hasta sus valores normales. Por otro lado, el descenso de la presión arterial por debajo de lo normal relaja los receptores de estiramiento y hace que el centro vasomotor se vuelva más activo de lo normal, provocando vasoconstricción y aumentando la acción de la bomba cardíaca.
Valores normales y características físicas de los principales componentes del líquido extracelular
Los valores fuera de estos intervalos suelen deberse a una enfermedad. Lo más importante es conocer los límites por encima de los cuales estas alteraciones provocan la muerte.
Características de los sistemas de control
Retroalimentación negativa de la mayoría de los sistemas de control: la mayoría de los sistemas de control actúan mediante ésta. Si algún factor se vuelve excesivo o deficiente, un sistema de control inicia una retroalimentación negativa, la cual se trata de una serie de cambios que devuelven ese factor hacia un determinado valor medio, manteniendo la homeostasis.
Ganancia de un sistema de control: la ganancia de una retroalimentación negativa es el grado de eficacia con el que un sistema de control mantiene sus condiciones constantes.
La retroalimentación positiva a veces provoca círculos viciosos y la muerte
Por lo general, la retroalimentación posivita llega a provocar una inestabilidad, misma que puede generar la muerte; se da cuando el estímulo inicial provoca más reacciones del mismo tipo, también es considerada como «círculo vicioso» . No obstante, en algunos casos puede llegar a ser útil, por ejemplo: en la coagulación sanguínea, el parto y la generación de señales nerviosas, ésta última permite que los nervios participen en los miles de sistemas de control de retroalimentación negativa de los nervios.
Siempre que la retroalimentación positiva es útil, la propia retroalimentación positiva forma parte de un proceso global de retroalimentación negativa.
Tipos más complejos de sistemas de control: control adaptativo
Control anterógrado: el cerebro lo aplica en los movimientos muy rápidos del organismo;este control hace que se contraigan los músculos apropiados, es decir, las señales del nervio sensible de las partes en movimiento informan al cerebro si el movimiento se está realizando correctamente. En caso contrario, el cerebro corrige las señales anterógradas que envía hacia los músculos la siguiente vez que se necesite ese movimiento. Y si necesita nuevas correcciones se realizarán cada vez en los movimientos sucesivos; es lo que se denomina control adaptativo.
LA CÉLULA Y SUS FUNCIONES
Organización de la célula
La célula se compone principalmente del núcleo y el citoplasma, separados entre sí por una membrana nuclear, también existe la membrana celular o plasmática, la cual separa el citoplasma de los líquidos circundantes.
Las diferentes sustancias que componen la célula se conocen colectivamente como protoplasma, compuesto por 5 sustancias fundamentales:
Agua: principal medio líquido, en una concentración de 70-85%. Muchos de los componentes químicos de la célula están disueltos, mientras que otros están en suspensión como micropartículas sólidas.
Iones: productos químicos inorgánicos de las reacciones celulares, necesarios para el funcionamiento de algunos de los mecanismos de control celulares. Algunos de los iones importantes de la célula son el potasio, el magnesio, el fosfato, el sulfato, el bicarbonato y cantidades más pequeñas de sodio, cloruro y calcio.
Proteínas: constituyen del 10 al 20% de la masa celular, hay 2 tipos:
Proteínas estructurales.- presentes en la célula en forma de filamentos largos que son polímeros de muchas moléculas proteicas individuales. Un uso importante de este tipo de filamentos intracelulares es la formación de microtúbulos que proporcionan los «citoesqueletos» de orgánulos celulares.
Proteínas funcionales.- compuestas por combinaciones de pocas moléculas en un formato tubular-globular. Son principalmente las enzimas de la célula y a menudo son móviles dentro del líquido celular. Además, muchas de ellas están adheridas a las estructuras membranosas dentro de la célula y las enzimas catalizan reacciones químicas al tener contacto directo con las demás sustancias presentes en el líquido.
Lípidos: constituyen el 2% de la masa de la célula, los de mayor importancia son los fosofolípidos y el colesterol, pues de ellos se forman las barreras de la membrana celular y de la intracelular. Algunas también tienen triglicéridos o grasas neutras, representan el principal almacén del organismo de nutrientes energéticos.
Hidratos de carbono: constituyen del 1 al 6% de la masa celular; tienen un papel importante en la nutrición celular y siempre están presentes en forma de glucosa disuelta en el líquido extracelular circundante, de forma que es fácilmente accesible a la célula, también se pueden almacenar en forma de glucógeno.
Estructura física de la célula
Estructuras membranosas de la célula
La mayoría de las membranas de orgánulos están formadas principalmente de lípidos, proteínas y enzimas, los primeros proporcionan una barrera que impide el movimiento de agua y sustancias hidrosolubles desde un compartimiento celular a otro; las proteínas suelen atravesar toda la membrana proporcionando vías especializadas que a menudo se organizan en poros auténticos para el paso de sustancias específicas a través de la membrana y las enzimas catalizan multitud de reacciones químicas diferentes.
Membrana celular o plasmática: es una estructura elástica, fina y flexible que tiene un grosor de 7,5 a 10 nm. Está formada mayormente por proteínas y lípidos, con una composición aproximada de:
55% de proteínas
25% de fosfolípidos
13% de colesterol
4% de otros lípidos
3% de hidratos de carbono
La barrera lipídica de la membrana celular impide la penetración del agua
Su estructura básica consiste en una bicapa lipídica, la cual es una película fina de doble capa de lípidos, cada una de las cuales contiene una sola molécula de grosor y rodea de forma continua toda la superficie celular. En ella se encuentran intercaladas grandes moléculas proteicas globulares. La bicapa lipídica básica está formada por moléculas de fosfolípidos. Un extremo de cada molécula de fosfolípido es soluble en agua, es decir, es hidrófilo (extremo fosfato del fosfolípido), mientras que el otro es soluble sólo en grasas (porción del ácido graso). La capa lipídica de la zona media de la membrana es impermeable a las sustancias hidrosolubles habituales, pero las sustancias hidrosolubles, como oxígeno, dióxido de carbono y alcohol pueden penetrar con facilidad .
Las moléculas de colesterol de la membrana también tienen una naturaleza lipídica, determinan el grado de permeabilidad de la bicapa y controlan gran parte de la fluidez de la membrana.
Proteínas de la membrana celular integrales y periféricas
Proteínas integrales: componen canales o poros a través de los cuales las moléculas de agua y las sustancias hidrosolubles pueden difundir entre los líquidos extracelular e intracelular, también tienen propiedades selectivas que permiten la difusión preferente de algunas sustancias específicas; actúan como proteínas transportadoras de sustancias; actúan como enzimas o como receptores, es decir, son un medio de transmisión de la información sobre el entorno hacia el interior de la célula.
Proteínas periféricas: se unen a las integrales, funcionan en su mayoría como enzimas o como controladores del transporte de sustancias a través de los poros de la membrana celular.
Hidratos de carbono de la membrana: «glucocáliz» celular
La mayoría de las proteínas integrales son glucoproteínas y casi la décima parte de las moléculas lipídicas de la membrana son glucolípidos. Las porciones «gluco» de estas moléculas casi siempre sobresalen hacia el exterior de la célula. Toda la superficie externa de la célula a menudo contiene un recubrimiento débil de hidratos de carbono que se conoce como glucocáliz, el cual tiene las siguientes funciones:
Tienen una carga eléctrica negativa que proporciona a la mayoría de las células una carga negativa a toda la superficie que repele a otros objetos negativos
El glucocáliz de algunas células se une al glucocáliz de otras, con lo que une las células entre sí
Actúan como componentes del receptor para la unión de hormonas
Algunas estructuras de hidratos de carbono participan en reacciones inmunitarias
Citoplasma y sus orgánulos
EL citosol es la parte del citoplasma en donde se dispersan los orgánulos, contiene principalmente proteínas, electrólitos y glucosa disueltos.
Retículo endoplásmico.- es una red de estructuras vesiculares tubulares y planas del citoplasma conectados entre sí; sus paredes también están formadas por membranas de bicapa lipídica que contienen grandes cantidades de proteínas. El espacio que queda dentro de los túbulos y vesículas está lleno de una matriz endoplásmica.
Ribosomas y retículo endoplásmico rugoso.- son partículas granulares diminutas; cuando están presentes, el retículo se denomina retículo endoplásmico rugoso. La función de los ribosomas es sintetizar nuevas moléculas proteicas en la célula.
Retículo endoplásmico agranular o liso.- es la parte del retículo endoplásmico que no tiene ribosomas; actúa en la síntesis de sustancias lipídicas y en otros procesos de las células que son promovidos por las enzimas intrarreticulares.
Aparato de Golgi.- está formado por cuatro o más capas apiladas de vesículas cerradas, finas y planas, que se alinean cerca de uno de los lados del núcleo. Este aparato es prominente en las células secretoras, donde se localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras. Funciona relacionado al retículo endoplásmico, ya que las sustancias atrapadas en las vesículas del RE se transportan desde el retículo endoplásmico hacia el aparato de Golgi.
Lisosomas.- son orgánulos vesiculares que se forman por la rotura del aparato de Golgi y después se dispersan por todo el citoplasma. Constituyen el aparato digestivo intracelular que permite que la célula digiera: Las estructuras celulares dañadas, las partículas de alimento que ha ingerido, las sustancias no deseadas; tiene un diámetro de 250 a 750 nm. Está rodeado por una membrana bicapa lipídica típica y está llena con grandes cantidades de gránulos pequeños, de 5 a 8 nm de diámetro.
Peroxisomas.- Se cree que están formados por autorreplicación y contienen oxidasas capaces de combinar el oxígeno con los iones hidrógeno para formar peróxido de hidrógeno, mismo que se utiliza junto a una catalasa para oxidar muchas sustancias que, de lo contrario, serían venenosas para la célula.
Mitocondrias.- También conocidos como centros neurálgicos de la célula, su número total varía de menos de 100 hasta varios miles. Se concentran en aquellas porciones de la célula que son responsables de la mayor parte de su metabolismo energético. Está compuesta por 2 membranas de bicapa lipídica-proteínas: una membrana externa y una interna. Además, la cavidad interna de la mitocondria está llena con una matriz que contiene grandes cantidades de enzimas disueltas que son necesarias para extraer la energía de los nutrientes. La energía liberada se usa para sintetizar una sustancia de «alta energía» que se denomina trifosfato de adenosina (ATP).
Citoesqueleto celular: estructuras filamentosas y tubulares
Las proteínas fibrilares de la célula se organizan habitualmente en filamentos o túbulos que se originan como moléculas proteicas precursoras sintetizadas por los ribosomas en el citoplasma. Las moléculas precursoras polimerizan después para formar filamentos.
Todas las células usan un tipo especial de filamento rígido formado por polímeros de tubulina para construir los microtúbulos, los cuales son estructuras tubulares fuertes.
Núcleo.- Centro de control de la célula, contiene la información genética, es decir, el ADN; se divide por medio de mitosis.
Membrana o cubierta nuclear.- son dos membranas bicapa separadas, una dentro de la otra. La membrana externa es una continuación del retículo endoplásmico del citoplasma celular y el espacio que queda entre las dos membranas nucleares también es una continuación con el espacio del interior del retículo endoplásmico. Varios miles de poros nucleares atraviesan la membrana nuclear.
Nucléolos y formación de ribosomas.- No tienen una membrana limitante, consisten en una acumulación simple de grandes cantidades de ARN y proteínas de los tipos encontrados en los ribosomas; aumenta de tamaño considerablemente cuando la célula está sintetizando proteínas activamente. La formación de los nucléolos y de los ribosomas comienza en el núcleo.
Sistemas funcionales de la célula
Endocitosis: es una función especializada de la membrana celular en donde las partículas muy grandes entran en la célula. Las formas principales de endocitosis son la pinocitosis y la fagocitosis.
La pinocitosis se refiere a la ingestión de partículas diminutas que forman vesículas de líquido extracelular y partículas dentro del citoplasma celular.
La fagocitosis se refiere a la ingestión de partículas grandes, como bacterias, células enteras o porciones de tejido degenerado.
Síntesis y formación de estructuras celulares en el retículo endoplásmico y el aparato de Golgi
Funciones específicas del retículo endoplásmico:
Las proteínas se forman en el retículo endoplásmico rugoso.
Síntesis de lípidos en el retículo endoplásmico liso
Proporciona las enzimas que controlan la escisión del glucógeno cuando se tiene que usar el glucógeno para energía
Proporciona una cantidad inmensa de enzimas que son capaces de detoxificar las sustancias
Procesamiento de las secreciones endoplásmicas en el aparato de Golgi: formación de vesículas
Extracción de energía de los nutrientes: función de la mitocondria
Las sustancias principales a partir de las cuales las células extraen energía son los alimentos, que reaccionan químicamente con el oxígeno: los hidratos de carbono, las grasas y las proteínas. En el cuerpo humano, esencialmente todos los hidratos de carbono se convierten en glucosa en el aparato digestivo y el hígado antes de que alcancen las demás células del organismo, al mismo tiempo las proteínas se convierten en aminoácidos y las grasas en ácidos grasos.
ATP
El ATP es un nucleótido compuesto por:
1) la base nitrogenada adenina
2) el azúcar pentosa ribosa
3) tres radicales fosfato.
Usos de ATP para las funciones celulares.
La energía del ATP se usa para promover tres categorías principales de funciones celulares:
1) transporte de sustancias a través de múltiples membranas en la célula
2) síntesis de compuestos químicos a través de la célula
3) trabajo mecánico
Locomoción de las células
Movimiento amebiano
Es el movimiento de toda la célula en relación con su entorno, como el movimiento de los leucocitos a través de los tejidos. Recibe su nombre por el movimiento de las amebas.
Tipos de células que muestran movimiento amebiano
Las células más frecuentes que muestran movimiento amebiano en el cuerpo humano son los leucocitos cuando salen de la sangre hacia los tejidos para formar macrófagos tisulares; los fibroblastos se mueven hacia una zona dañada para reparar el daño e incluso las células germinales de la piel se desplazan hacia la zona de un corte para reparar el desgarro.
La locomoción celular es especialmente importante en el desarrollo del embrión y el feto después de la fertilización de un óvulo.
Cilios y movimientos ciliares
Un segundo tipo de movimiento celular, el movimiento ciliar, es un movimiento a modo de látigo de los cilios que se encuentran en la superficie de las células. Este movimiento existe sólo en dos lugares del cuerpo humano: en la superficie de las vías respiratorias y en la superficie interna de las trompas uterinas (trompas de Falopio) del aparato reproductor.
Cada cilio es una excrecencia de una estructura que se apoya inmediatamente por debajo de la membrana celular, el cuerpo basal del cilio.
El flagelo de un espermatozoide es similar a un cilio; de hecho, tiene el mismo tipo de estructura y el mismo tipo de mecanismo contráctil. Sin embargo, este flagelo es mucho más largo y se desplaza con ondas de tipo cuasi-sinusoidal en lugar de movimientos de tipo látigo.
Esta es tu primera entrada. Haz clic en el enlace «Editar» para modificarla o eliminarla, o bien crea una entrada. Si lo prefieres, puedes utilizar esta entrada para contar a los lectores por qué has empezado este blog y qué tienes previsto hacer con él. Si necesitas ayuda, contacta con los simpáticos miembros de los foros de ayuda.

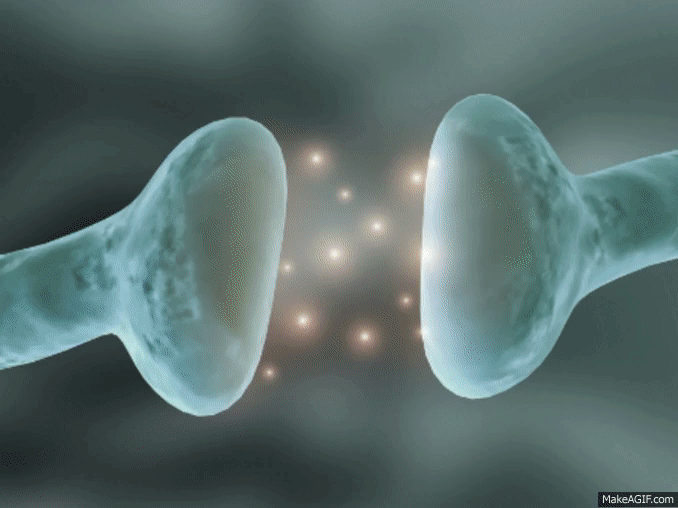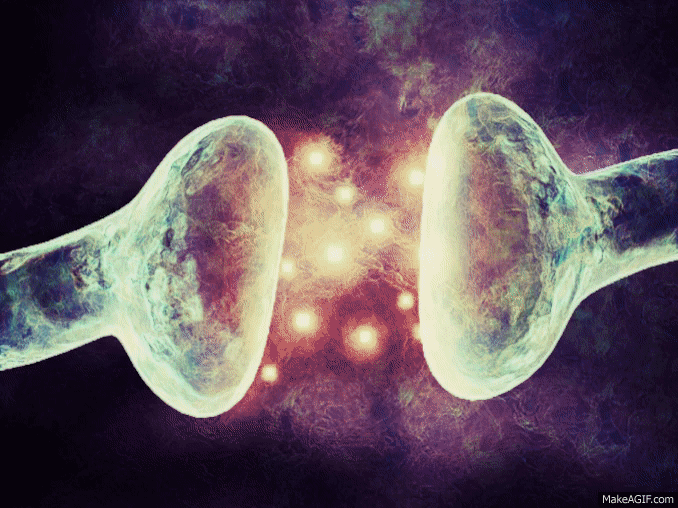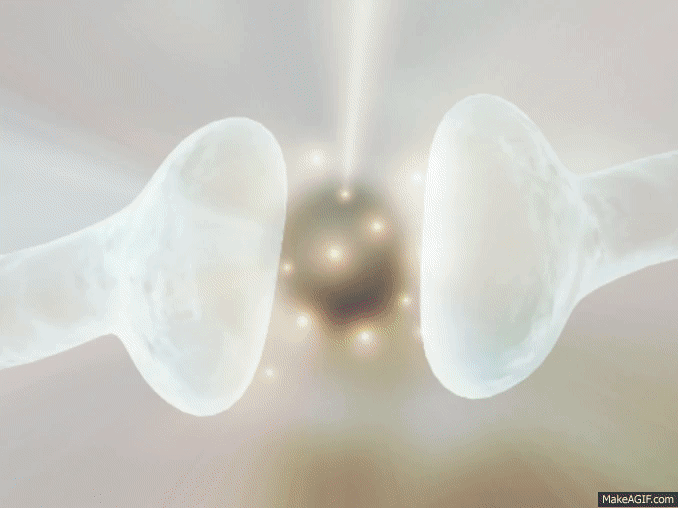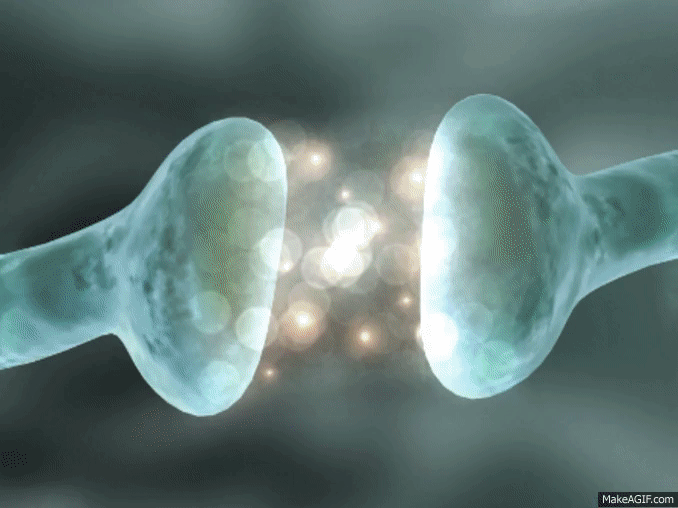


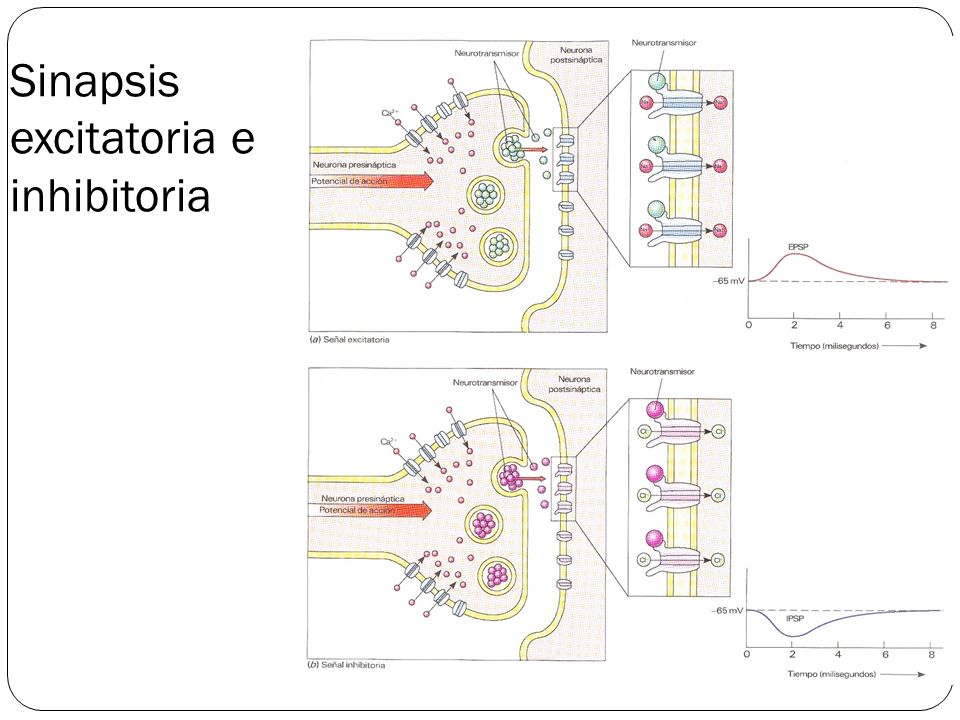
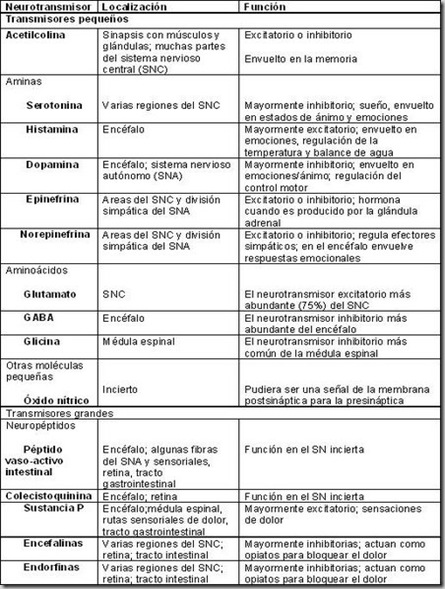
 El corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre hacia los órganos periféricos. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Los ventrículos aportan la principal fuerza del bombeo que impulsa la sangre.
El corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre hacia los órganos periféricos. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Los ventrículos aportan la principal fuerza del bombeo que impulsa la sangre.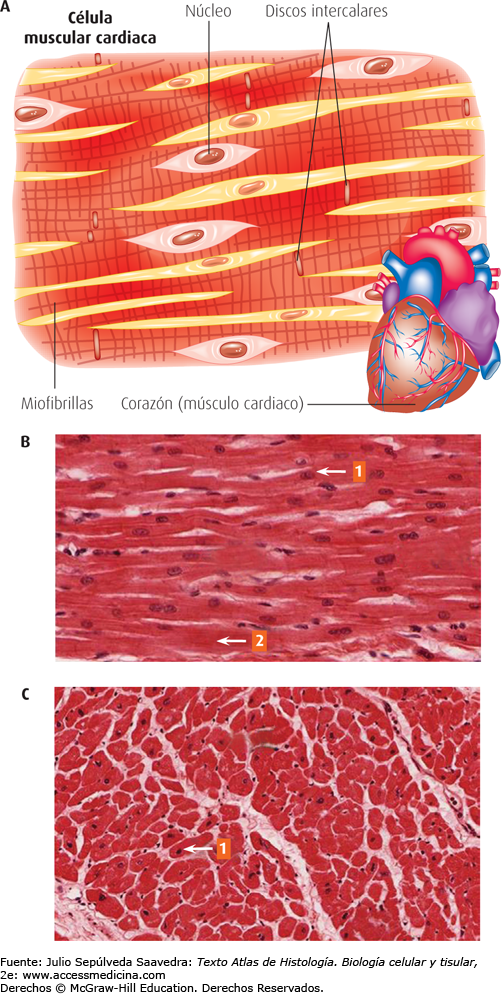 El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. La duración de la conducción es mucho mayor y las fibras contienen pocas fibrillas contráctiles y presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón.
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. La duración de la conducción es mucho mayor y las fibras contienen pocas fibrillas contráctiles y presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón.


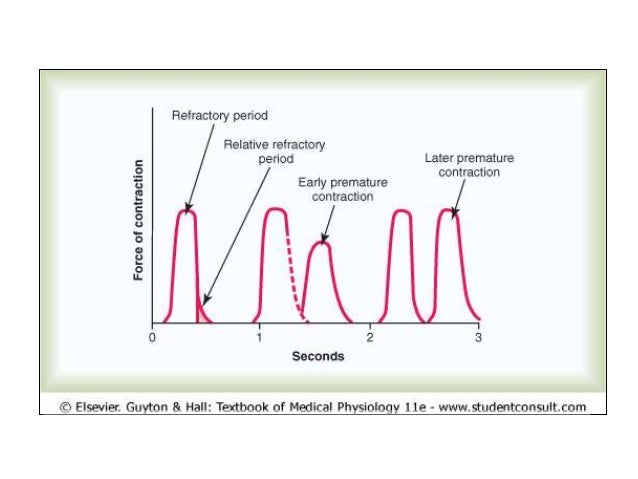

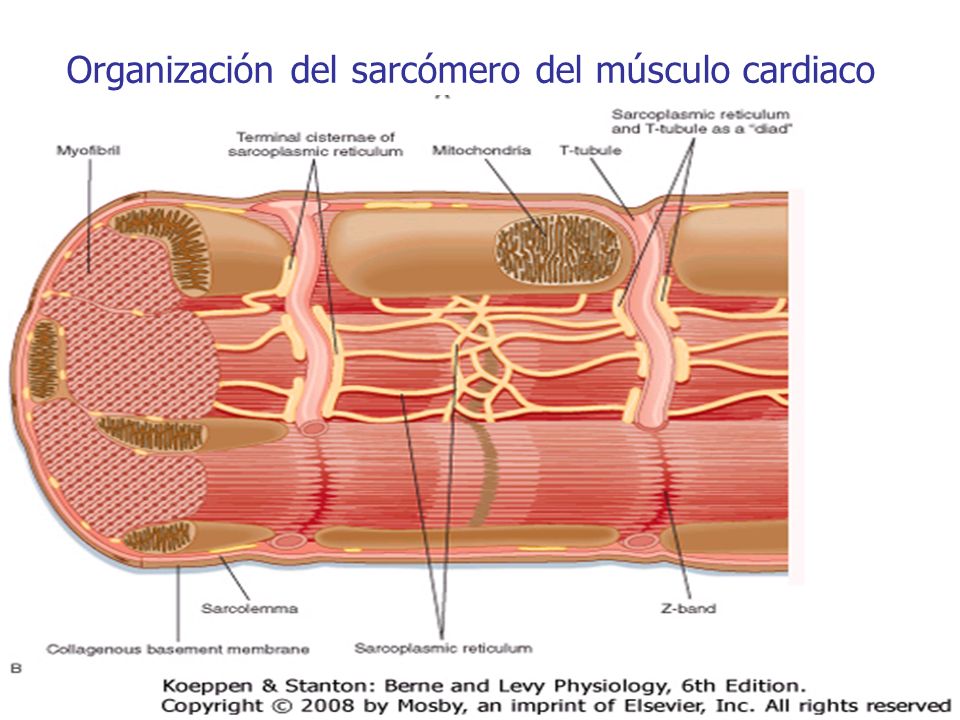
 Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la gene ración espontánea de un potencial de acción en el nódulo sinusal que está localizado en la pared superolateral de la aurícula derecha. El potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Las aurículas se contraen antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular. Por tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.
Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco. Cada ciclo es iniciado por la gene ración espontánea de un potencial de acción en el nódulo sinusal que está localizado en la pared superolateral de la aurícula derecha. El potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Las aurículas se contraen antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular. Por tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.


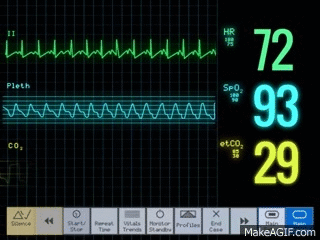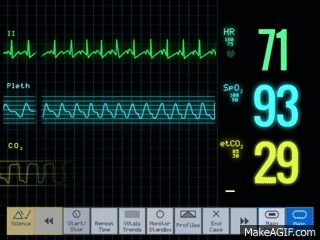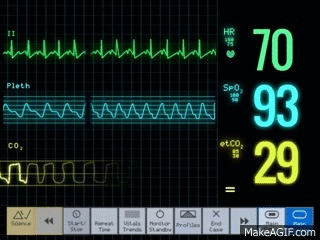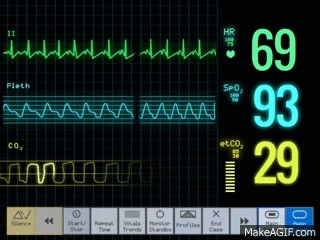


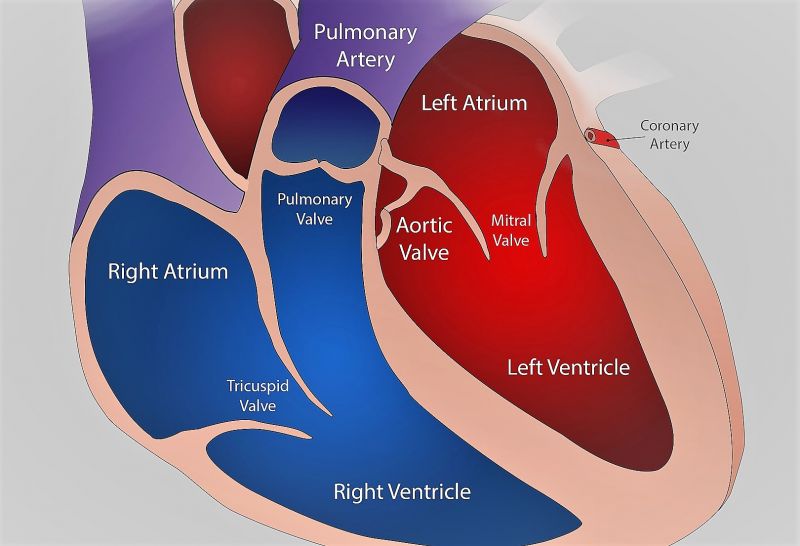




 El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1min. El trabajo del corazón se utiliza para mover la sangre desde las venas de baja presión hacia las arterias de alta presión. Esto se denomina trabajo volumen-presión o trabajo externo. Además, una pequeña proporción de la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar. Este es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco.
El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1min. El trabajo del corazón se utiliza para mover la sangre desde las venas de baja presión hacia las arterias de alta presión. Esto se denomina trabajo volumen-presión o trabajo externo. Además, una pequeña proporción de la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar. Este es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco.


 El consumo de oxígeno ha demostrado ser también casi proporcional a la tensión que se produce en el músculo cardíaco durante la contracción multiplicada por la duración de tiempo durante la cual persiste la contracción, denominada índice de tensión-tiempo. Se gasta mucha más energía química a presiones sistólicas normales cuando el ventrículo está dilatado anómalamente debido a que la tensión del músculo cardíaco durante la contracción es proporcional a la presión multiplicada por el diámetro del ventrículo.
El consumo de oxígeno ha demostrado ser también casi proporcional a la tensión que se produce en el músculo cardíaco durante la contracción multiplicada por la duración de tiempo durante la cual persiste la contracción, denominada índice de tensión-tiempo. Se gasta mucha más energía química a presiones sistólicas normales cuando el ventrículo está dilatado anómalamente debido a que la tensión del músculo cardíaco durante la contracción es proporcional a la presión multiplicada por el diámetro del ventrículo.







 Masa de de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra y hacer que las fibras musculares se contraigan simultáneamente.
Masa de de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra y hacer que las fibras musculares se contraigan simultáneamente.


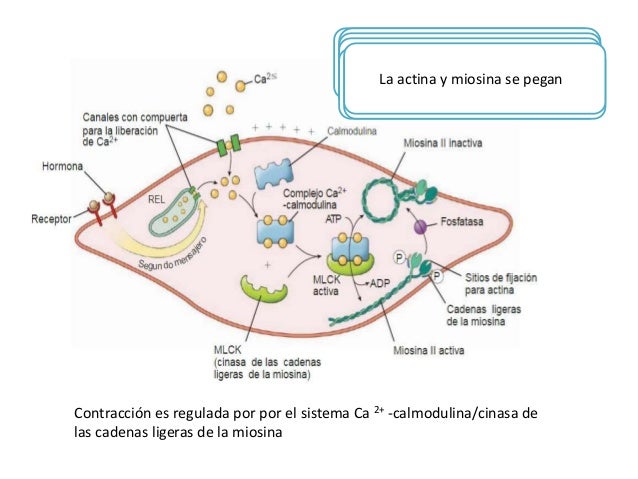




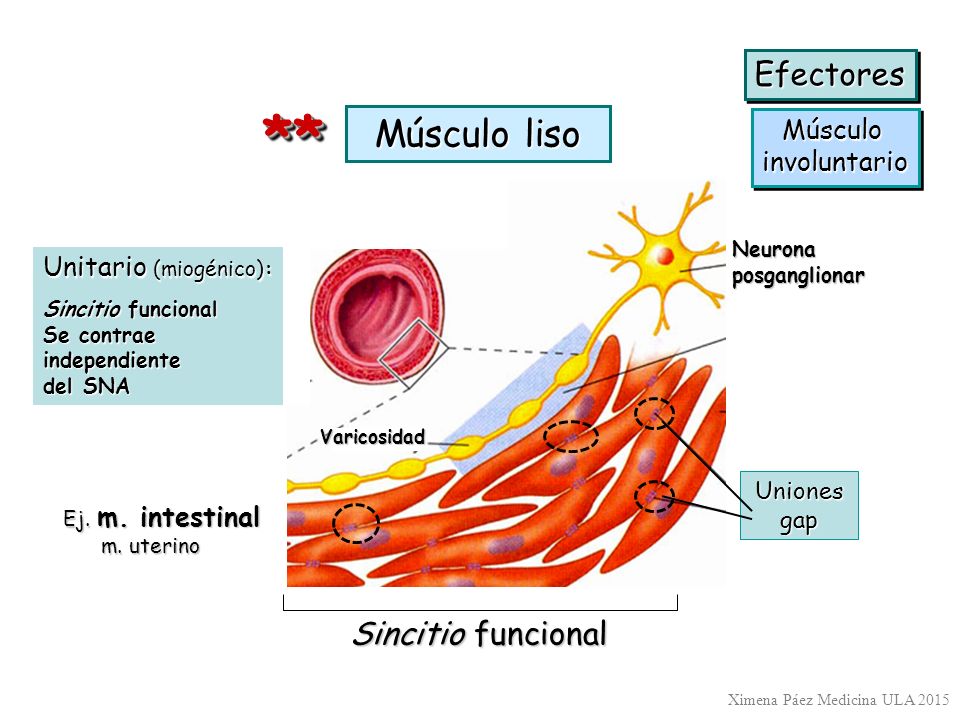

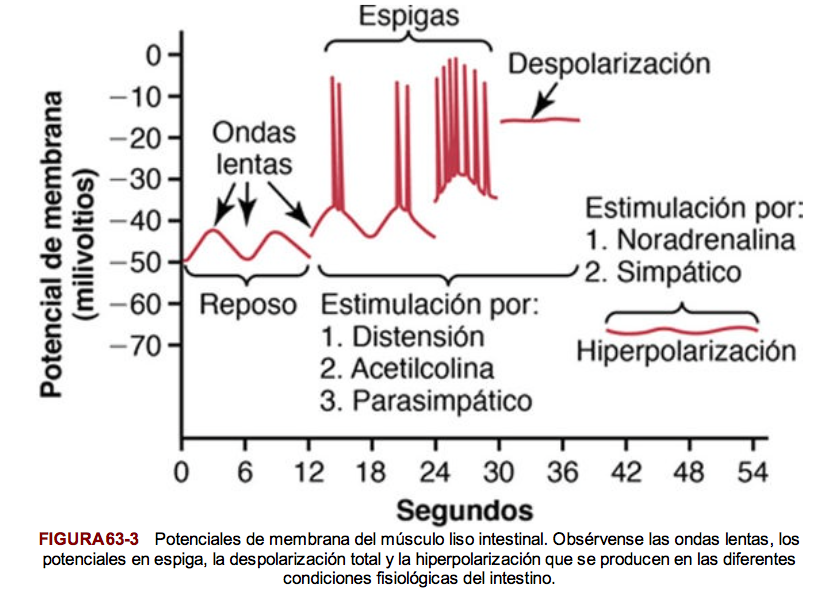

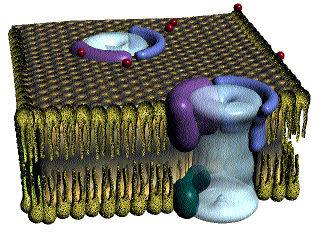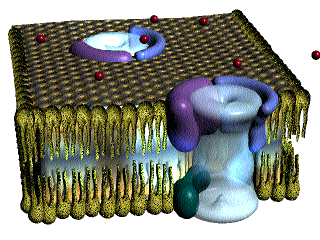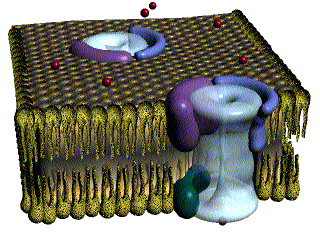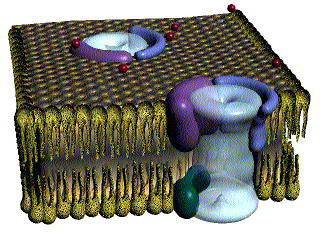




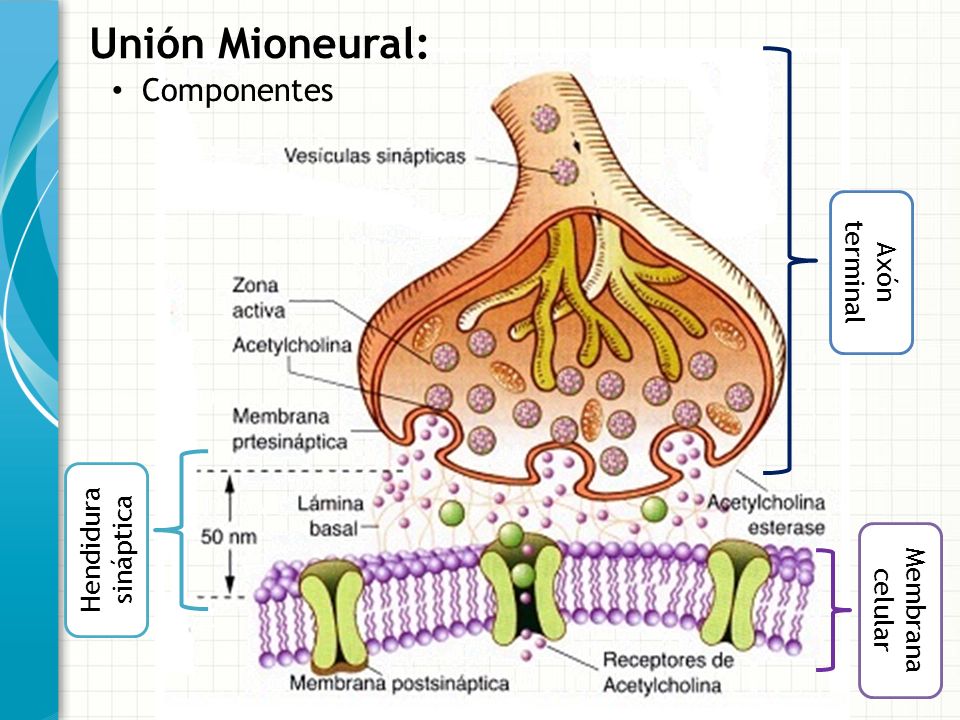
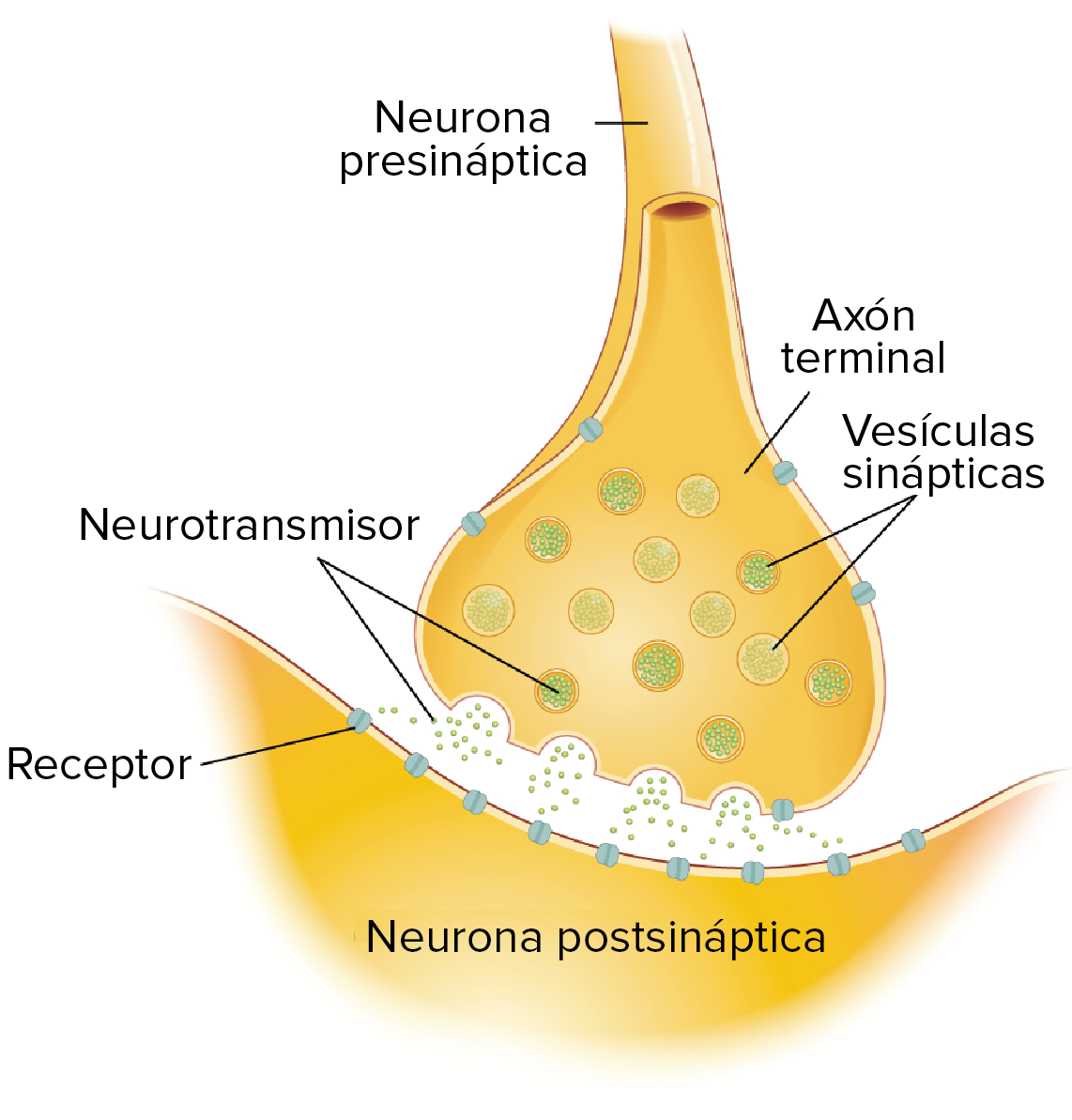
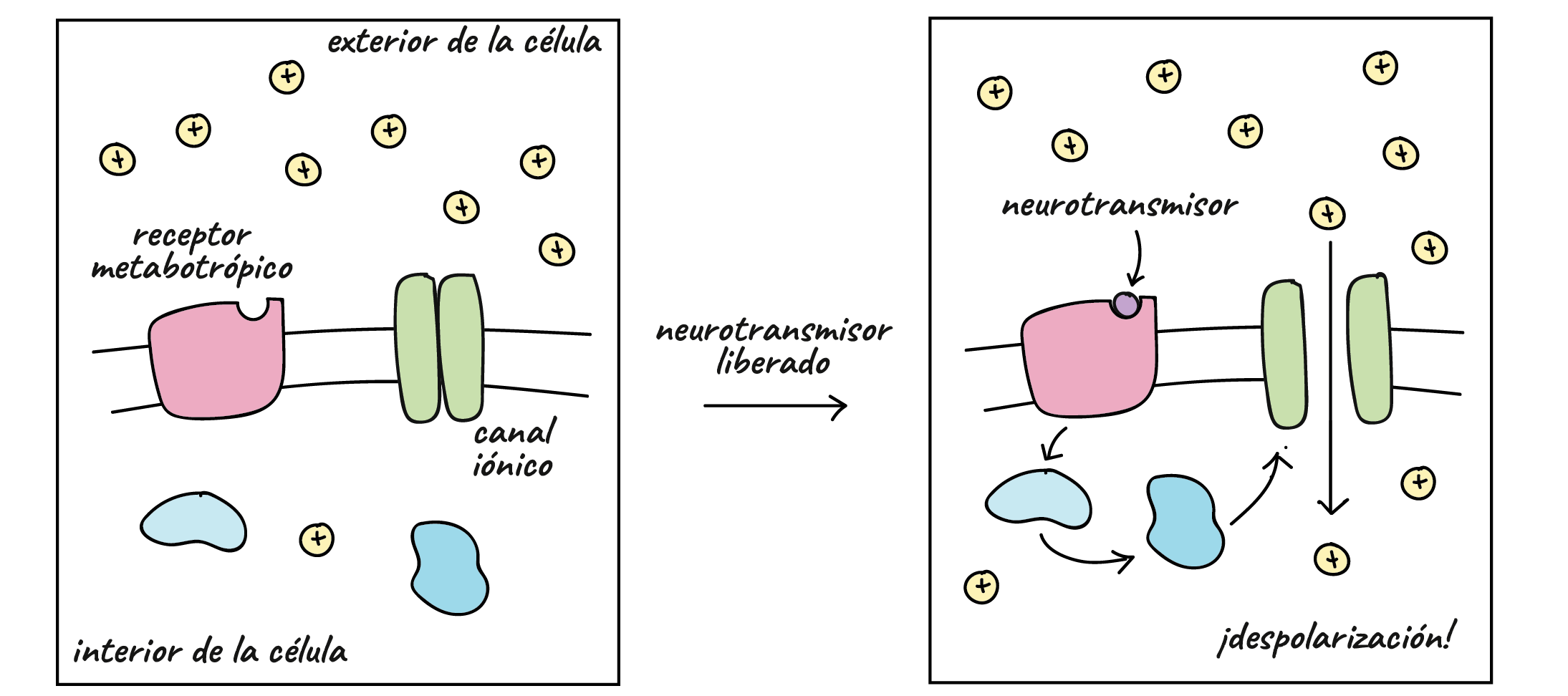

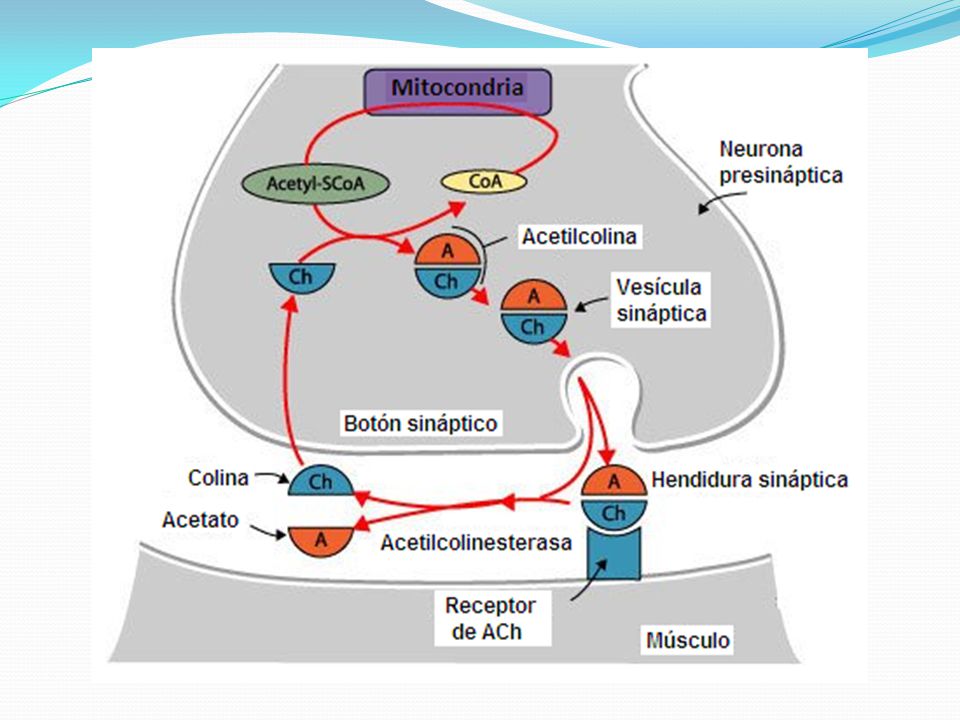





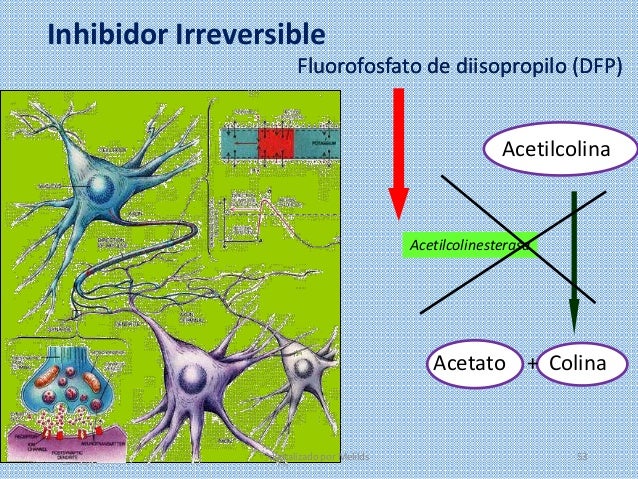

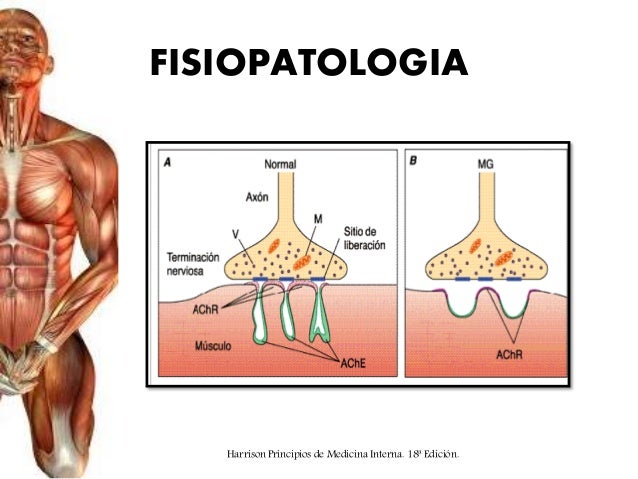 La miastenia grave produce parálisis muscular debido a que las uniones neuromusculares no pueden transmitir suficientes señales desde las fibras nerviosas a las fibras musculares. En cuanto a su patogenia, en la sangre se han detectado anticuerpos dirigidos frente a los receptores de acetilcolina.
La miastenia grave produce parálisis muscular debido a que las uniones neuromusculares no pueden transmitir suficientes señales desde las fibras nerviosas a las fibras musculares. En cuanto a su patogenia, en la sangre se han detectado anticuerpos dirigidos frente a los receptores de acetilcolina.
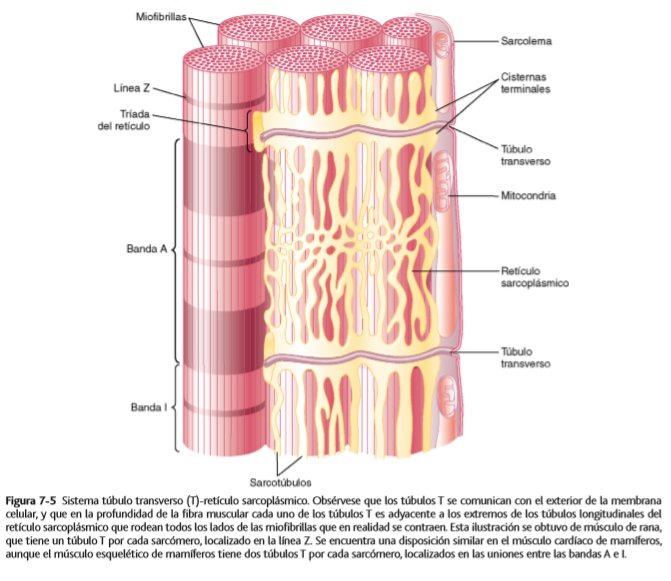
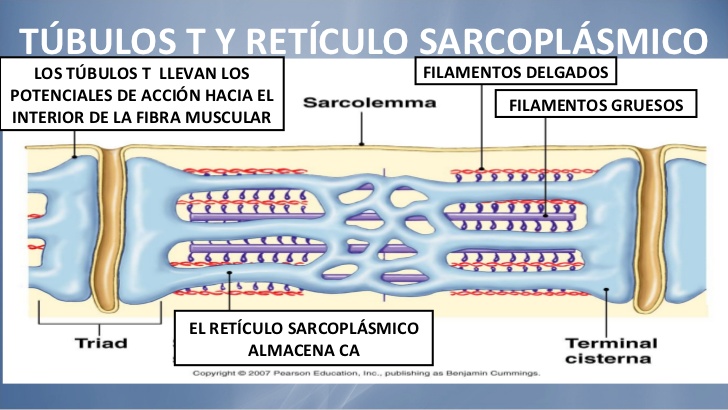

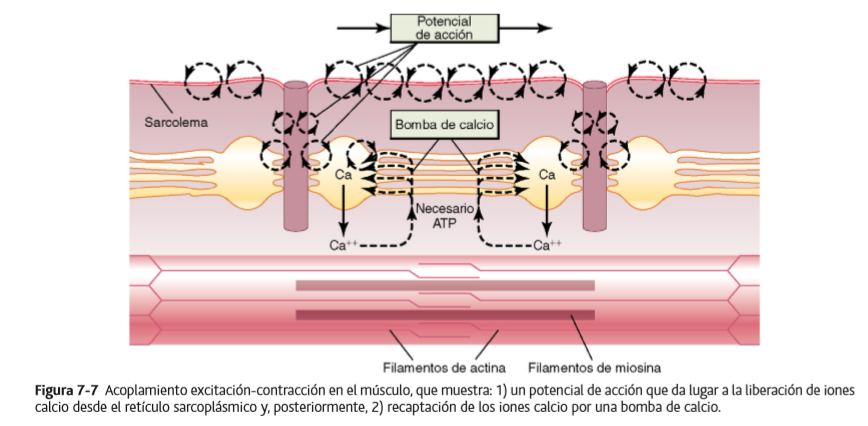



































 Hay muchos tipos diferentes de músculo; algunos son largos y se contraen una distancia larga, y algunos son cortos pero tienen áreas transversales grandes y pueden proporcionar una fuerza de contracción extrema en distancias pequeñas. El estudio de los diferentes tipos de músculos, de los sistemas de palanca y de sus movimientos se denomina cinesiología.
Hay muchos tipos diferentes de músculo; algunos son largos y se contraen una distancia larga, y algunos son cortos pero tienen áreas transversales grandes y pueden proporcionar una fuerza de contracción extrema en distancias pequeñas. El estudio de los diferentes tipos de músculos, de los sistemas de palanca y de sus movimientos se denomina cinesiología. 




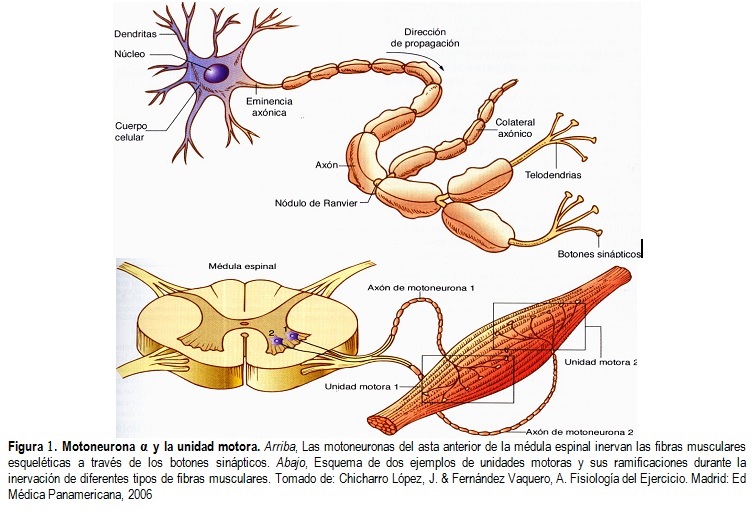












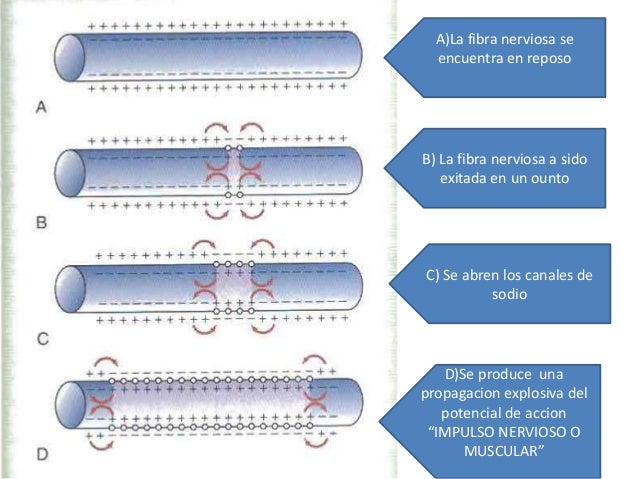











 La membrana celular contiene grandes números de moléculas proteicas insertadas en los lípidos . La bicapa lipídica constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular, aunque hay unas sustancias que pueden penetrarla y difundirse directamente a través de la propia sustancia lipídica. La mayoría de las proteínas penetrantes actúan de formas diferentes. Por ejemplo, las proteínas de los canales y las transportadoras, las primeras permiten el movimiento libre de agua, iones o moléculas a través de la membrana, mientras que las segundas se unen a ellos para transportarlos; sin embargo, ambas son muy selectivas para los tipos de molecúlas o iones que pueden atravesar la membrana.
La membrana celular contiene grandes números de moléculas proteicas insertadas en los lípidos . La bicapa lipídica constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular, aunque hay unas sustancias que pueden penetrarla y difundirse directamente a través de la propia sustancia lipídica. La mayoría de las proteínas penetrantes actúan de formas diferentes. Por ejemplo, las proteínas de los canales y las transportadoras, las primeras permiten el movimiento libre de agua, iones o moléculas a través de la membrana, mientras que las segundas se unen a ellos para transportarlos; sin embargo, ambas son muy selectivas para los tipos de molecúlas o iones que pueden atravesar la membrana.



















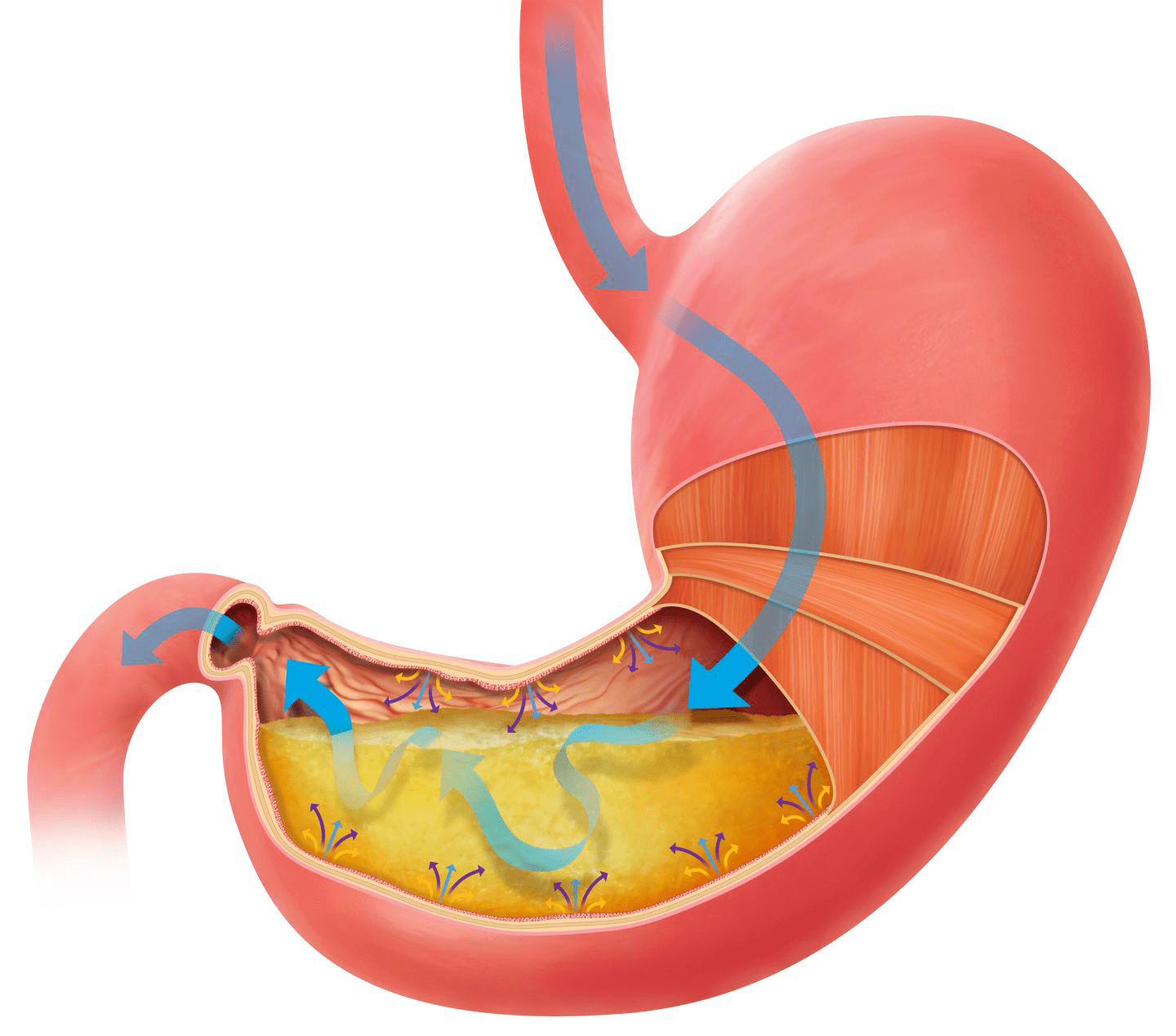
 colectores corticales de los riñones, donde hay células intercaladas especiales. Aquí es donde se secretan grandes cantidades de iones hidrógeno desde la sangre hacia la orina con el objetivo de eliminar de los líquidos corporales el exceso de iones hidrógeno.
colectores corticales de los riñones, donde hay células intercaladas especiales. Aquí es donde se secretan grandes cantidades de iones hidrógeno desde la sangre hacia la orina con el objetivo de eliminar de los líquidos corporales el exceso de iones hidrógeno.











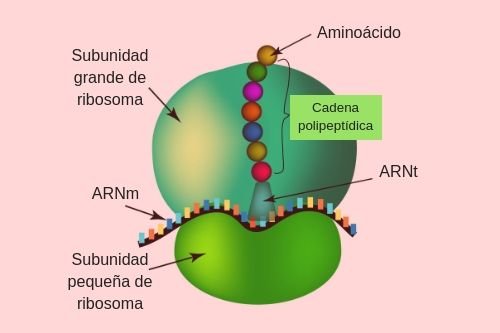

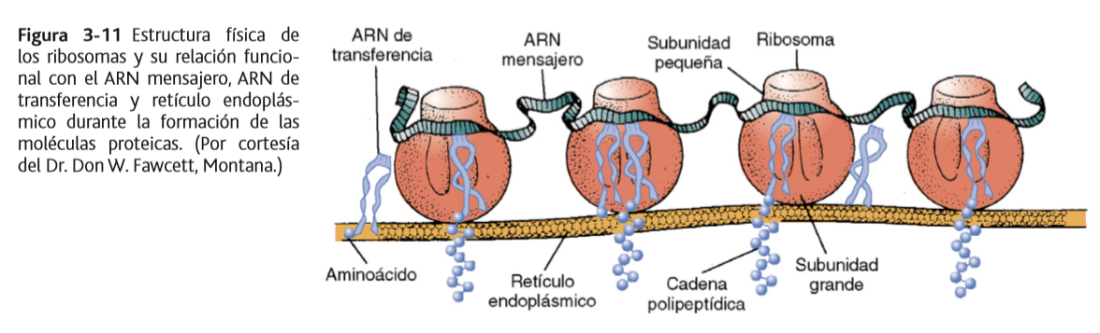





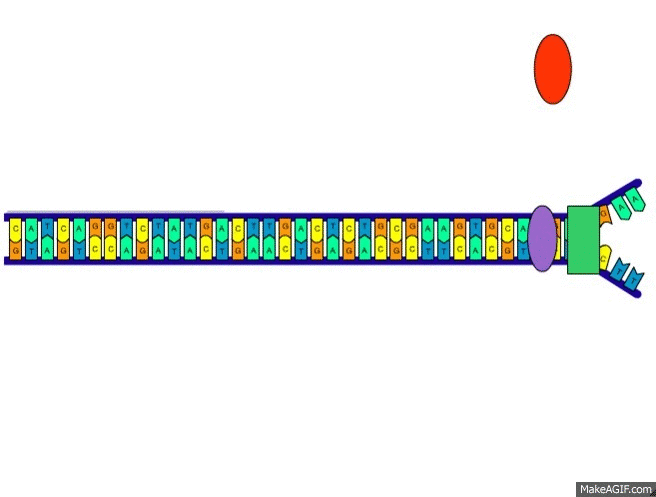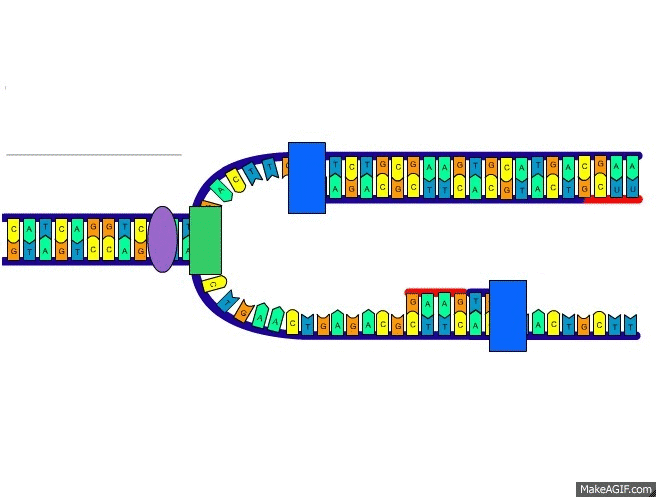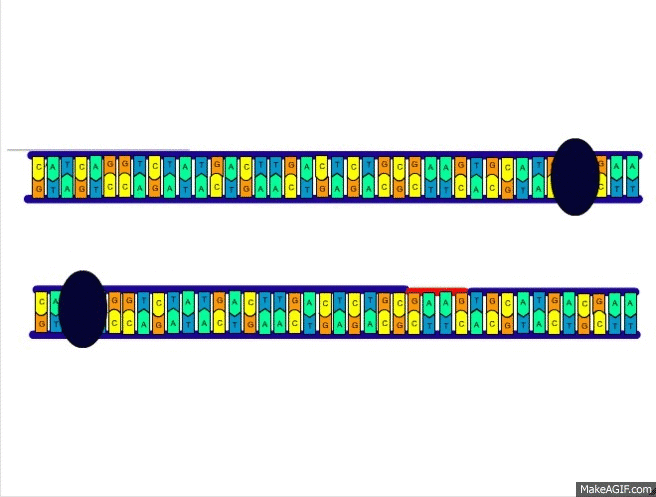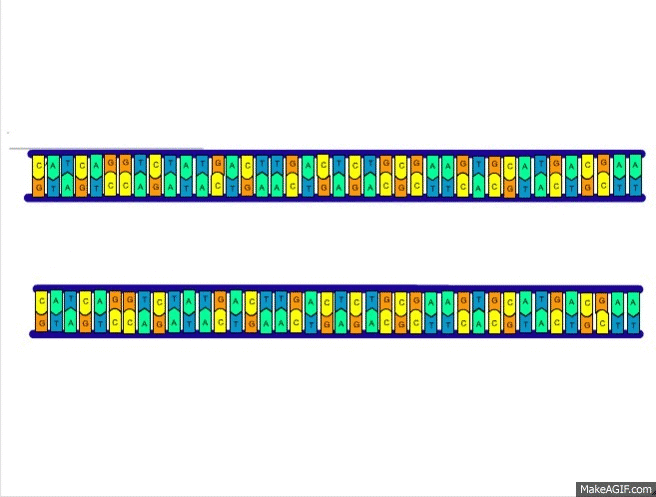



 Aparato mitótico: función de los centriolos
Aparato mitótico: función de los centriolos






 Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis.
Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis.

 Se debe en todos o casi todos los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los genes anormales se denominan oncogenes y se han descu bierto hasta 100 tipos distintos., aunque también hay antioncogenes, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer.
Se debe en todos o casi todos los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los genes anormales se denominan oncogenes y se han descu bierto hasta 100 tipos distintos., aunque también hay antioncogenes, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer.